Les résultats sont présentés sous forme de moyenne ± SE et, selon les cas, les valeurs individuelles minimales et maximales ont été précisées [m - M].
III . RESULTATS
Masse corporelle et hibernation :
Chez la marmotte adulte, les évolutions de la masse corporelle du mâle et de la femelle (Fig. l) montrent des variations saisonnières nettement marquées et parallèles, la masse corporelle des femelles étant légèrement inférieure à celle des mâles.
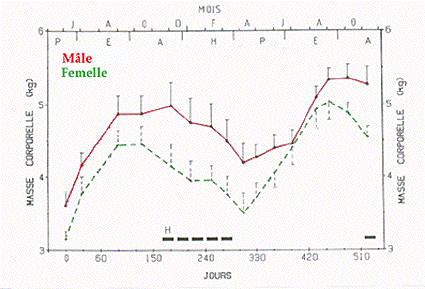
Figure 1 - Evolutions saisonnières simultanées de la masse corporelle chez la Marmotte adulte mâle (-*-) et femelle (-*-). Moyennes ± SE ; H = hibernation.
Chez le mâle, l'évolution de la masse corporelle est caractérisée par : des valeurs maximales en automne (de sept. = 4,88 ± 0,25 kg [4,28 - 5,76] à début déc. = 4,98 ± 0,32 kg [4,28 - 6,15], une décroissance régulière en hiver jusqu'à un minimum en avril (avril = 4,19 ± 0,27 kg [3,24 - 4,80] et une augmentation lente au printemps puis rapide en été (de début juil. = 4,46 ± 0,19 kg à sept. - oct. = 5,35 ± 0,19 kg [4,36 - 5,71]).
Chez la femelle, le profil de l'évolution saisonnière de la masse corporelle est comparable, avec un maximum plus court en fin d'été - début d'automne (sept. - oct. = 4,45 ± 0,25 kg [3,95 - 4,71], un minimum aussi début avril (3,51 ± 0,27 kg [3,13 - 4,03]) puis une croissance régulière jusqu'à un maximum en septembre (5,04 ± 0,25 kg [4,56 - 5,37]).
Au cours de l'année, l'amaigrissement hivernal varie, chez les mâles et les femelles, respectivement de 15 à 21% et l'engraissement estival de 27 à 43%. La période d'hibernation, déterminées par la mesure de la température corporelle lors des prélèvements sanguins, a été observée de novembre à mi-mars dans nos conditions climatiques.
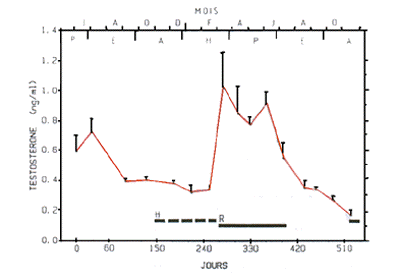 Figure 2 - Evolution saisonnière de la testostéronémie chez la Marmotte mâle adulte. Moyennes ± SE ;
Figure 2 - Evolution saisonnière de la testostéronémie chez la Marmotte mâle adulte. Moyennes ± SE ;
H = hibernation ; R = testicules descendus dans le scrotum.
Reproduction :
Chez le mâle, les testicules, intraabdominaux pendant la majeure partie de l'année, descendent dans le scrotum à partir de mi-mars - début avril, et l'estimation du volume testiculaire par palpation est alors possible. Ainsi, les testicules sont très gros en avril et leur volume décroît lentement jusqu'en juillet. En août, ils sont de nouveau en position intraabdominale. Les concentrations plasmatiques de la testostérone (T) chez la marmotte montrent des variations de faible amplitude (valeurs minimales : 0,2-0,3 ng/ml) ; valeurs maximales : 1,1-1,8 ng/ml). L'activité endocrine testiculaire présente cependant une évolution saisonnière nette (Fig. 2) caractérisée par : une phase de repos d'août à février (T < 0,4 ng/ml), une reprise rapide entre février (0,34 ± 0,03 ng/ml) et mars (1,3 ± 0,23 ng/ml), des concentrations maximales de mars à juin (T > 0,8 ng/ml), et une décroissance au début de l'été (juillet = 0,55 ± 0,11 ng/ml) aboutissant dès août (0,35 ± 0,05 ng/ml) au repos sexuel.
Chez la femelle, au cours de l'année, les concentrations circulantes d'oestradiol ont toujours été inférieures aux limites minimales de sensibilité de notre dosage. Pour la progestérone, le dosage a permis de détecter des variations uniformes et de faible amplitude (0,2-0,4 ng/ml) tout au long de l'année, sans relation avec un état physiologique ou une activité saisonnière particulière. Il est aussi à noter que dans nos conditions d'élevage aucune gestation n'a pu âtre observée et, qu'à ce jour, nous manquons de références convenables puisque aucune analyse n'a pu âtre effectuée chez des femelles gestantes.
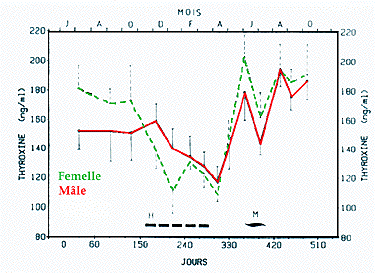 Figure 3 - Evolutions saisonnières simultanées de la thyroxinémie chez la Marmotte adulte mâle (-*-) et femelle (-*-). Moyennes ± SE ; H = hibernation ;
Figure 3 - Evolutions saisonnières simultanées de la thyroxinémie chez la Marmotte adulte mâle (-*-) et femelle (-*-). Moyennes ± SE ; H = hibernation ;
M = mue.
Fonction thyroïdienne :
La détermination des concentrations plasmatiques de la thyroxine (T4) a montré dans les conditions de notre dosage (RIA Sigma) de larges variations entre 100 et 200-220 ng/ml. Dans les deux sexes l'évolution saisonnière de la T4 est comparable (Fig. 3) et caractérisée par : des concentrations élevées (140-160 ng/ml) de juin à octobre (femelles) ou décembre (mâles), une décroissance en hiver et des valeurs minimales en avril (110 ng/ml), une réactivation printanière avec un premier maximum en juin (180 ng/ml). Dans les deux sexes, une décroissance est observée en juillet (140 ng/ml), en mâme temps que s'effectue la mue. Pendant l'hibernation, chez les animaux en hypothermie, nous avons pu trouver indifféremment des valeurs de T4 faibles ou très élevées.
IV . DISCUSSION ET CONCLUSIONS
Chez la marmotte alpine, maintenue dans les conditions climatiques naturelles de la forât de Chizé, des cycles saisonniers marqués continuent d'âtre observés. Ces variations annuelles des fonctionnements endocriniens ne paraissent pas montrer de différences sensibles avec les observations disponibles actuellement pour cette espèce dans son habitat alpin naturel. La différence essentielle nous parait âtre liée à la durée et, surtout, à l'intensité de l'hibernation (facteurs dont les paramètres n'ont pas été suffisamment étudiés au cours de ce travail). Cependant, en dépit de la captivité, le maintien de l'hibernation et d'une rythmicité annuelle marquée des paramètres étudiés permet de considérer la présente étude comme un travail de base nécessaire et indispensable à la compréhension de l'adaptation de l'animal à son milieu.
Comme chez tous les hibernants, la diminution de la masse corporelle s'effectue principalement pendant la période d'hibernation. Chez la femelle, la diminution plus rapide de la masse corporelle en début d'automne pourrait peut-âtre s'expliquer par un jeûne spontané associé ou non à une activité générale encore élevée, Chez la marmotte l'engraissement s'effectue par anticipation au cours de l'été (de juin - juillet à fin août début septembre). Chez le mâle, probablement en relation avec les conditions d'élevage et le climat local, la masse corporelle se maintient élevée en plateau au cours de l'automne. Au cours de la vie active, les valeurs minimales de la masse corporelle sont concomitantes de la phase d'activité testiculaire printanière élevée et, au cours de l'été, l'engraissement parait facilité par l'involution de la testostéronémie. Chez le mâles, la réactivation de la fonction gonadique s'effectue rapidement à la sortie de l'hibernation et l'activité endocrine testiculaire est de courte durée (mars à juin-début juillet). Dans les deux sexes, les taux de thyroxine les plus bas sont bien corrélés avec l'amaigrissement des animaux et concomitants de la période d'hibernation. Chez la marmotte mâle, contrairement à ce qui a été observé chez un autre hibernant : le Hérisson (Saboureau & Boissin, 1983), la testostéronémie et la thyroxinémie évoluent en opposition de phase, sauf en mai-juin, ce qui témoigne de la possibilité de cycles indépendants et/ou de régulations centrales différentes.
Chez la marmotte femelle, l'absence de variation de l'oestradiol et de la progestérone peut âtre liée aux conditions de manipulation des animaux et, en particulier, à l'anesthésie. Cependant, il n'est pas impossible de penser que chez cette espèce sauvage l'ovulation puisse âtre provoquée, comme chez Marmota monax (Sinha Hikim et al., 1992), ce qui expliquerait l'absence de cycle détectable.
En conclusion, cette étude a montré chez la marmotte alpine l'existence de variations endocrines saisonnières nettes dans les deux sexes. La confrontation de ces premiers résultats avec des déterminations obtenues chez les animaux vivant en altitude devrait nous permettre de valider ces évolutions saisonnières et (ou) de préciser certains points. Ces données endocrinologiques sont aussi intéressantes à approfondir, en relation avec les éthologistes, pour la compréhension des comportements observés pendant la vie active au sein du groupe social.
BIBLIOGRAPHIE
Berges R. (198l). Cycles saisonniers de la fonction testiculaire chez deux hibernants, le lérot (Eliomys quercinus L.) et le spermophile (Citellus tridecemlineatus). Leur régulation par la température ambiante. Thèse de doctorat de 3e cycle, Université de Toulouse, France.
Boissin J. & Canguilhem B. (1988). Les rythmes circannuels chez les mammifères. Arch. Int. Physiol. Biochim., 96 : A289-A345.
Canguilhem B. (1985). Rythmes circannuels chez les mammifères hibernants sauvages. Can. J. Zool. 63 : 453-463.
Canguilhem B., Vivien-Roels B., Demeneix B., Miro J.L., Masson-Pevet M. & Pevet P. (1986). Seasonal and endogenous variations of plasma testosterone (T), thyroxine (T4), triiodothyronine (T3) in the European hamster. In Endocrine regulations as adaptive mechanisms to the environment, pp. 29l-295, Eds I. Assenmacher & J.Boissin, Editions du C.N.R.S., Paris.
EI Omari B., Lacroix A. & Saboureau M. (1989). Daily and seasonal variations in plasma LH and testosterone concentrations in the adult male hedgehog (Erinaceus europaeus L.). J. Reprod. Fert., 86 : 145-155.
Jallageas M. & Assenmacher I. (1983) Annual plasma testosterone and thyroxine cycles in relation to hibernation in the edible dormouse Glis glis. Gen. Comp. Endocrinol., 50 : 452-467.
Saboureau M. & Boissin J. (1983). Endocrine cycles and hibernation in the hedgehog. In Mechanisms of adaptation to natural variations in the environment. In Adaptations to terrestrial environments, pp. 203-217, Eds N. S. Margaris, M. Arianoutsou-Farraggitaki & R. J. Reiter, PIenum Publishing Corporation, New York.
Saboureau M., Maurel D. & Boissin J. (1986). Behavioural and endocrine adaptations to the seasonal environmental variations in two wild mammals: the hedgehog and the European badger. In Endocrine regulations as adaptive mechnanisms to the environment, pp. 35l-36l, Eds I. Assenmacher & J. Boissin, Editions du C.N.R.S., Paris.
Sinha Hikim A. P., Woolf A., Bartke A. & Amador A. G. (1992). Further observations on estrus and ovulation in woodchucks (Marmota monax) in captivity. Biol. Reprod., 46 : l0-16.
Remerciements
Ce travail a été réalisé grâce au soutien financier du Ministère de la Recherche (contrat CNRS, Programme Environnement). Il a pu se dérouler grâce au soutien actif de l'administration et des agents du Parc National de la Vanoise , que nous tenons à remercier.
A suivre...
Retour ou sommaire