MARMOTS ARE ZOONOSIS PROVOKING CARRIERS
Â
Bibikov D.I.
Abstract - Marmots are widely spread in open landscapes of North Hemisphere Mountains and plains. Within their range they mosaicly disperse and have low density. Large size and big shelters-burrow are responsible for the major role they play in biocoenosis and in conservation of natural foci-zoonosis.
Peculiarities in marmot ecology and behaviour create conditions favourable for microbe and virus transmission. A list of marmot diseases and the pattern of distribution, microbe circulation and activity of natural foci in marmots is here presented and discussed.
Marmots are widely spread in open landscapes of North Hemisphere Mountains and plains. Within their range they mosaicly disperse and have low density (up to 30 and rarely 300 individuals/km2). Large size and big shelters - burrows, are responsible for the major role they play in biocoenosis, and in conservation of natural foci-zoonosis.
Peculiarities in marmot ecology and behaviour create conditions favourable for microbe and virus transmission.
The list of marmot diseases is rather long, but incomplete (Table 1).
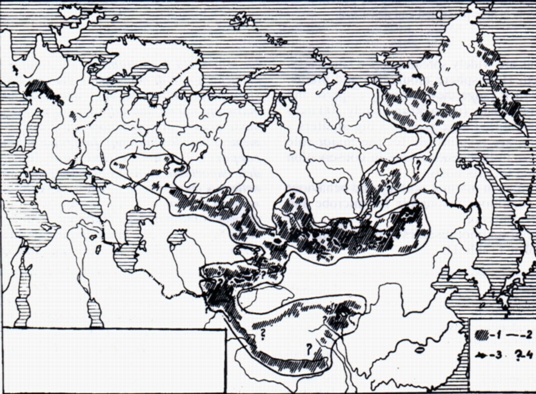
Rather often epizootics of several diseases (e.g. different combinations rabbit-fever, pseudotuberculosis, listeriosis, plague) occur in the same area. Most zoonoses in marmots haven't been well studies. We have detail descriptions only for the pasteurellosis, Powassan encephalitis, and in particular for the plague. Thus, we consider the plague in the Middle and Central Asia to analyse the patterns of distribution, microbe circulation and activity natural foci in marmots. Natural foci of the plague cover a smaller part of the marmot range in the Transbaikal Area, Mongolia, China, Tien-Shan, Alai, Gissar Mountains (Fig. 1).
The Marmota camtschatica, M. bobac schaganensis, M. b. bobac and M. marmota ranges are outside these foci. The isolation of the plague microbe from M. flaviventris or their fleas is known in some areas of the USA and British Columbia, Canada.
Except warm-blooding marmots, Biocoenotic structure of the Asiatic plague foci includes different Cittelus, Ochotona, Microtus etc. species. In the eastern foci, in the Transbaikal area and Mongolia, other rodents, except major carriers-marmots, play important role. In Tien-Shan these foci are nearly monohostal foci. Marmot fleas Oropsylla silantiewi, Rhadinopsylla liventricosa, Ceratophyillus lebedowi, C. dolabris etc. transmit a plague microbe from a rodent-donor to a healthy recipient. In wintertime, the plague preserves in marmot and fleas, which survive in abandoned burrows without food and in permanent low temperature condition more than 420 days.
Spatial structure of the plague enzootic area is characterised with autonomous foci, isolated from each other for centuries by marmot range breaks (Fig. l). The combination of different habitats: which are constantly optimum for rodents and their fleas life-meadow-steppes and nonstable dry steppes and alpine meadows is necessary. Droughts in dry steppes and overhumidity in alpine meadows are responsible for sharp decrease in forage.
In both types of foci the lack of food increases marmot movements, and interpopulation interactions which create more favourable conditions for a plague epizootic (Bibikov, 1965; Bibikov et al., 1973).
The smallest portions of these structures are plague microfoci, which includes different in their conditions marmot colonies, where epizootics exist for years moving from one family to another.
POPULATION FACTORS
Marmots are very sensitive to the plague microbe when are active, and resistant to it over hibernation period or before it. Thus, of 108 M. baibacina experimentally infected in August, 23 animals (21 percent) died before hibernation started, in 7 other animals infection generalization was observed in spring, the rest survived (Peisakhis, 1958). Resistance to infection in animals of different sex and age, and fatness varies greatly. Death from plague in some families decreases numbers and changes family structure, numbers of ill individual in a family may reach 22 percent. At the same time, movements of the survived animals to look for a pair grow; and abandoned burrows become contemporary shelters for marmot-migrants, which attract plague flea. Eco-physiological heterogeneity and changes in population structure during epizootics are responsible for family epizootic movements within plague foci and their long existence.
Seasonal and annual changes in epizootic activity depend on shifts in population structure and dynamics, and animal mobility caused by weather and food condition variations, In early spring, marmots are well-fed and less sensitive to the plague microbe and attached to winter burrows. An epizootic does not develop until they move to summer burrows where plague fleas have preserved. The aggravation occurs in mid-summer when animals are active and their resistance to the plague is low. As for as hibernation in coming, animal fatness grows and the same does their resistance to the microbe. The interpopulation contacts are getting lower, and epizootics are going out slowly. In eastern foci epizootics go on in autumn either, because pikas and voles are involved.
Favourable weather conditions provoke marmot increase in rather unsuitable habitats. And, when a weather cycle changes migrations might increase, animal physiological conditions might get worse, so a plague epizootic is activated.
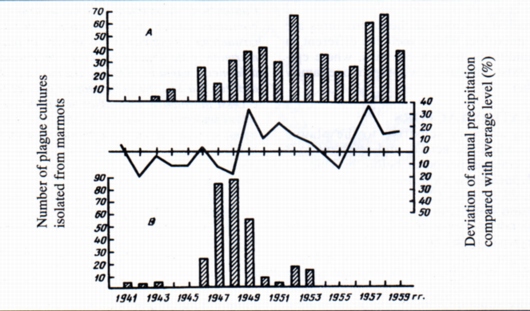
Evidently, conditions for epizootics to occur, are different in arid high mountains or alpine meadow (Fig. 2 A, B).
In landscapes with cold weather conditions and high humidity, epizootics occurred after periods with low summer temperatures and surplus rainfall (1949-1954 and 1957-1959). On the contrary, in arid high mountains, epizootics happened in 1946-1949, which followed several dry summer seasons (Fig. 2B).
Sanitation measures in the Tien-Shan foci in 1950-60 covered marmot number decreasing which made microbe transmission more difficult. The foci were not eliminated, however, epizootic intensity sharply reduced, some of them died away.
Observation revealed ecological mechanisms, which preserve microbes in decreased marmot populations: 1. rearrangement of survived single animals in optimum habitats and transmission of infected fleas; 2. slowed-down epizootic development in preserved microfoci; 3- long-term existence of foci supported by the plague microbe from the population of additional carriers-susliks, pikes and voles.
Thus, observations on plague focus sanitation and their consequent restoration showed the importance of ecologo-geographical factors for natural focus zoonosis development.
ritorno/back
LA MARMOTTA COME VEICOLO DI ZOONOSI
Bibikov D.I.
La Marmotta è ampiamente distribuita negli ambienti aperti di montagna e pianura dell'emisfero nord. All'interno degli areali occupati le marmotte hanno una diffusione a mosaico con basse densità di popolazione (fino a 30 e raramente 300 individui/Kmq). Le grandi dimensioni corporee e le ampie tane sono le ragioni dell'importante ruolo svolto da questi animali nelle biocenosi e nella conservazione dei focolai naturali di zoonosi. Le caratteristiche ecologiche e comportamentali della Marmotta creano condizioni favorevoli alla trasmissione di batteri e virus. L'elenco delle malattie che colpiscono questa specie è piuttosto lungo, ma incompleto (Tabella 1). Spesso eventi a carattere epizootico di diverse malattie (Febbre del Coniglio, Pseudotubercolosi, Listeriosi, Peste) avvengono contemporaneamente nella stessa area. La maggior parte delle zoonosi della Marmotta non sono state studiate approfonditamente. Possediamo descrizioni accurate solo della Pasteurellosi, dell'Encefalite di Powassan, ed in particolare della Peste.Prendiamo quindi in considerazione quest'ultima malattia, presente in Asia centrale, per analizzare le caratteristiche di distribuzione, di diffusione microbica e di attività naturale dei focolai di Peste nella Marmotta. I focolai naturali di Peste coprono un'area ristretta dell'areale distributivo della Marmotta nella regione del Transbaikal, in Mongolia, Cina, Thien-Shan, Alai e nelle montagne di Gissar (Fig. 1). Gli areali di Marmota camtschatica, M. bobac schaganensis, M. b. bobac e M. marmota sono al di fuori di questi focolai.à altresi noto l'isolamento del bacillo della Peste da M. flaviventris o da loro ecto-parassiti (pulci) in alcune aree degli USA e della Colombia britannica, in Canada. Oltre alla Marmotta la struttura biocenotica dei focolai asiatici di Peste include diverse specie di Cittelus, Ochotona, Microtus, ed altri micro-mammiferi. Nei focolai orientali, nel Transbaikal e in Mongolia, altri Roditori, oltre alla Marmotta, giocano un ruolo importante. Nel Thien-Shan i focolai presentano quasi sempre un solo ospite. Le pulci delle marmotte (Oropsylla silantiewi, Rhadinopsylla liventricosa, Ceratophyllus lebedowi, C. dolabris, ecc.) trasmettono l'agente della Peste da un roditore-donatore ad uno sano-ricevente. Durante l'inverno la Peste si mantiene all'interno dell'organismo della marmotta ed in quello degli ectoparassiti.Questi ultimi sono in grado di sopravvivere nelle tane abbandonate, senza cibo e in condizioni di basse temperature costanti per più di 420 giorni.
Distribuzione spaziale
La distribuzione spaziale dei casi di infezione di Peste enzootica è caratterizzata da focolai autonomi, separati gli uni dagli altri da migliaia di anni, a seguito di soluzioni di continuo nell'areale di distribuzione della Marmotta (Fig. l). Per l'attivazione di un focolaio è necessaria la concomitante presenza di diversi habitat: alcuni costantemente ottimali per la sopravvivenza delle marmotte e delle pulci (praterie-steppe) ed altri a idoneità variabile (steppe asciutte e praterie alpine). La siccità nelle steppe asciutte e gli eccessi di umidità nelle praterie alpine causano infatti improvvise carenze di cibo, che aumentano gli spostamenti delle marmotte e le interazioni tra le diverse popolazioni. Tali eventi creano condizioni favorevoli per l'instaurarsi di un'epizoozia di Peste (Bibikov, 1965; Bibikov et al., 1973). Le forme più ristrette di distribuzione della malattia sono rappresentate da microfocolai di Peste, che interessano colonie in condizioni molto diverse, in cui l'infezione rimane per anni, spostandosi da un nucleo famigliare ad un'altro.
Fattori di popolazione
Le marmotte sono molto sensibili all'agente della Peste durante la fase di vita attiva, resistenti prima e durante l'ibernazione. Di 108 individui di M. baibacina infettati sperimentalmente in agosto, 23 (21%) morirono prima dell'ibernazione, 7 mostrarono la generalizzazione della malattia nella primavera successiva, mentre le rimanenti sopravvissero (Peisakhis, 1958).La resistenza all'infezione in animali di sesso, età e grado di ingrassamento diversi varia notevolmente. La morte per Peste riduce il numero di individui e modifica la struttura delle famiglie e la percentuale di individui malati in una famiglia può raggiungere il 22%. Nel contempo aumentano i movimenti degli individui sopravvissuti alla ricerca di un partner e le tane abbandonate possono fungere da rifugi temporanei per marmotte migratrici, che cosi si infestano di pulci infette. L'eterogeneità eco-fisiologica e i cambiamenti nella struttura di una popolazione che avvengono durante le epizoozie sono responsabili degli interscambi fra famiglie e quindi della lunga persistenza dei focolai di Peste.
Cambiamenti annuali e stagionali
Le variazioni annuali e stagionali nell'andamento delle epizoozie dipendono dai cambiamenti della struttura, della dinamica delle popolazioni e della motilità degli individui legati alle diverse disponibilità di cibo ed ai fattori climatici. All'inizio della primavera le marmotte sono ben nutrite, ancora legate all'uso delle tane invernali e quindi meno sensibili al bacillo della Peste. La malattia non si sviluppa finché gli animali non si spostano nelle tane estive dove le pulci della Peste sono sopravvissute. La situazione si aggrava a metà del periodo estivo, quando gli animali sono attivi e quindi la loro resistenza alla Peste si abbassa. All'approssimarsi del letargo l'animale accumula grasso ed aumenta nel contempo la resistenza al bacillo. I contatti tra le popolazioni si riducono e le epizoozie regrediscono lentamente. Nei focolai nei settori più orientali le epizoozie continuano anche in autunno, in quanto vengono interessati dalla malattia anche i Lagomorfi ed i piccoli Roditori. Condizioni climatiche favorevoli portano ad un incremento delle marmotte anche in aree generalmente poco idonee e, a seguito del peggioramento del clima, i fenomeni migratori possono aumentare, le condizioni fisiche degli animali peggiorano e l'infezione si diffonde. Ovviamente le condizioni predisponenti che favoriscono l'insorgere di una epizoozia in aree aride montane o nette praterie alpine sono differenti (Fig. 2 A, B). In ambienti caratterizzati da clima freddo e umido le epizoozie avvengono dopo estati fredde e più piovose della media (1949-1954 e 1957-1959). Al contrario, nelle montagne alte ed aride, l'epizoozia di Peste ebbe luogo nel periodo 1946-1949, a seguito di più stagioni estive molto secche (Fig. 2 B). Le misure sanitarie intraprese nei focolai nel 1950-1960 nel Tien-Shan comportarono la riduzione del numero delle marmotte rendendo più difficile la trasmissione dell'infezione. I focolai non vennero eliminati ma l'intensità del fenomeno epizootico si ridusse nettamente, anche se alcune marmotte morirono comunque.Le osservazioni rivelarono i meccanismi attraverso i quali gli agenti eziologici si mantenevano anche in popolazioni di dimensioni ridotte: 1) Ridistribuzione degli animali solitari sopravvissuti verso aree migliori e trasmissione delle pulci infette; 2) Rallentamento dello sviluppo della epizoozia nei rimanenti microfocolai; 3) Persistenza a lungo periodo dei focolai di Peste grazie alla presenza di bacilli provenienti da altre specie animali vettrici, quali susliks, pikas e arvicole. Queste osservazioni sugli interventi sanitari nei focolai di Peste e sulla loro ricomparsa dimostrano l'importanza dei fattori ecologici e geografici nello sviluppo naturale dei focolai di zoonosi.
ritorno/back
Â
ANANIN V.V. Leptospirosis in people and animals. M.: Medecine Publishers, 1971. 350 p.
BIBIKOV D.I. Marmota Frisch genus, 1775 - marmots. In: Medical Terriology. Ed. Kucheruk V. V. M.: Nauka Publishers, 1979, 269-279.
BIBIKOV D.I., BERENDYAEV S.A. et al. Natural foci of marmot plague. M.: Medicine Publishers, 1973, 192 p.
KALYAKIN V.N. Toxoplasmosis in mammals. In: Annual report. Medical Encyclopedia, 1971, vol. 3, 865-904.
KARULIN B.E., PCHELKINA A.A. Warm-blooding animals - carriers of mite spotted fever in Asia North. Sci. papers. USSR Academy of Sciences, 1958, vol. 120, N°1, 223-224.
KUCHERUK V.V., RYUTIN V.A., DUNAYEVA T.N. Study experience on Pasteurellosis epizootic of the Mongolian bobak in Mongolia East. In: Fauna and ecology of rodents, issue 4. Moscow Society of Naturalists,
1951, 82-97.
PEISAKHIS L.A. On pathogenesis of plague in grey marmots. Release 1 and 2. Sci. papers of the Middle Asia anti-plague Inst., Alma-Ata, 1958, vol. 4, 81.
RALl Yu.M. Rodents and natural foci of plague. M.: Medicine Publishers, 1960, 224 p.
TIMOFEYEVA L.A., GOLOVACHEVA V.Ya. Plague microbe and other microflora receiving in the Transbaikal plague locus. Sci. Papers. Irkutsk Anti-plague Inst. Chita, 1961, vol. 2, 105-106.
FYODOROVA N.l. Epidimiology and preventive measures against Ku-rikketsiosus. M.: Medicine Publishers, 1968, 251 p.
SHAMOVA A.M. A case of plague and pseudotuberculosis culture receiving from rodents in an enzootic focus of plague. Bull. Irkutsk Anti-plague Inst. of Siberia and Far East. 1959, vol. 21, 63-67.
BIBIKOV D.I. Spatial laws of natural focality of plague in Marmots. Proceed. of Symposium Theoretical Questions of natural Foci of Diseases (ed. B. Rosicky, K. Heyberger). Czechosl. Acad. of Sci., 1965, p. 83-88.
MCLEAN D.M., BEST J.B. et al. Powassan virus: summer infection cycle. - Canad. Med. Assoc. J., 1964, vol. 91. n° 26, p. 1360-1362.
MCLEAN D.M., SMITL P.A. et al. Powassan virus: vernal spread during 1965 - Canad. Med. Assoc. J., 1966, vol. 94, n° 11, p. 532-536.
POLLITZER R. Plague. Genève, 1954, p. 698.