Bassano B., Durio P., Gallo Orsi U., Macchi E
eds., 1992. Proceedings of the 1st Int. Symp. on
Alpine Marmot and genera Marmota. Torino.
Edition électronique, Ramousse R., International Marmot Network, Lyon 2002
ENERGETICS OF HIBERNATING AND NORMOTHERMIC ALPINE MARMOTS
Ortmann S. and Heldmaier G.
FB Biologie/zoologie, Philipps-Universitat, D-3550 Marburg
Abstract - Alpine marmots show a seasonal acclimatization of physiological parameters to lower the costs for thermoregulation and to support fattening in fall. The basal metabolic rate is reduced to 63 % of the spring value and the body temperature of normothermic marmots decreased within the thermoneutral zone. This decrease was more pronounced in fall than in spring and summer. The regulation of Tb to lower temperatures allowed the extension of the TNZ by shifting Tlc to lower values. Hibernation reduced energetic costs to 5.8 % of winter BMR of a normothermic marmot.
INTRODUCTION
Alpine marmots are confronted with strong seasonal variations in ambient temperature and food supply. As a means of energy conservation and an adaptation to cold and snow that covers the vegetation they spend the winter months in deep hibernation, huddling together in groups of up to twenty individuals in the depth of their burrows. Animals that show hibernation need to store energy reserves to comply with the physiological requirements of existence either by gathering food in their burrows or by accumulating body fat. Marmots exclusively rely on fat stores as the fuel for the hibernating season that last approximately six months. The remaining six months of the year are time of reproduction and intense and sustained energy intake accompanied by body mass production.
Body weight gain requires that energy intake exceeds energy expenditure and mammals show a variety of adaptations that minimizes energy output e.g. reduced aboveground activity or acclimatization of metabolic rate. The aim of this study was to investigate the extent of seasonal acclimatization of thermoregulatory energy demands and we focussed our attention on the regulation of body temperature and metabolic rate throughout the year.
METHODS
The marmots in our laboratory were kept in an outdoor enclosure provided with an indoor nesting box and fed ad lib with a mixture of grains, fresh grass and water. In order to measure the metabolic rate (VO2) animals were transferred into a Plexiglas box provided with a small layer of hay and placed in a climate chamber while normothermic or a walk-in climate room during hibernation respectively. Oxygen consumption and carbon-dioxide production were measured in an open system (for methodological details see Heldmaier and Steinlechner, 1981), body temperature (Tb) was recorded using temperature sensitive transmitters and a computerized recording system. Ambient temperature (Ta), measured by a thermocouple attached to the wall of the Plexiglas box, ranged from -0.5°C to 15°C during hibernation and from -50°C to 30°C during normothermia. VO2, Tb and Ta at a given ambient temperature were determined in six minute intervals and measured for at least 24h during hibernation or in one minute intervals for at least one hour while animals were normotherm. The minimal VO2 was calculated by averaging the three lowest values each temperature step.
The hibernation experiments were performed with seven individuals in winter 1988/89 and 1989/90, measurements of metabolic rate during normothermia with four animals in April, July, October and December 1990.
RESULTS
1.Seasonal changes of Tb and VO2
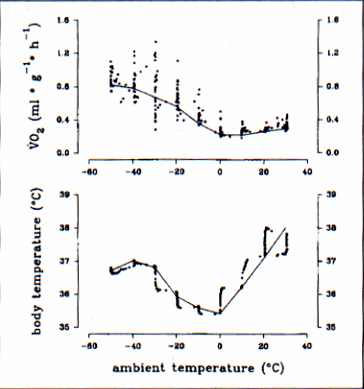
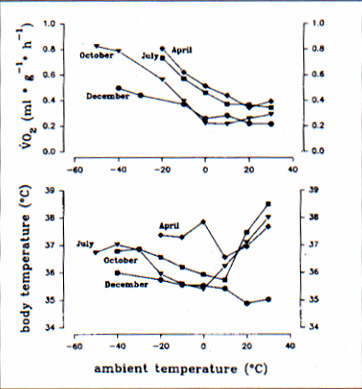
Body temperature and oxygen consumption at ambient temperatures between 30°C and -50°C of a single normothermic marmot measured in October are illustrated in figure 1. Both parameters did not remain constant throughout cold exposure. Metabolic rate remained minimal for Ta between 30°C and 0°C and increased linearly with a further decrease in ambient temperature. Body temperature reached a maximum at Ta 30°C, decreased to a minimal value of 35.4°C and increased again at ambient temperatures below 0°C. Weight specific resting metabolic rate and body temperature at various ambient temperatures were measured at different times of the year and the results of one representative individual are presented in figure 2. We found a shift in the thermoneutral zone (TNZ) to lower values throughout the year as well as a seasonal rhythm of basal metabolic rate (BMR) with high values in spring and midsummer and low values in fall and winter. The animals body weight ranged between 2915 g in spring and 4220 g in October.
Within this span there was no obvious relation between mass specific VO2 and body mass. During the year lower critical temperature T1c was reduced from 20°C in spring to 0°C in fall and increased again to 20°C in winter. This reduction of basal metabolic rate in fall and winter lowers the energetic costs of single Alpine marmots in the thermoneutral zone to 63 % of the value a marmot has to invest in spring or summer. As mentioned above body temperature did not remain constant at various ambient temperatures. The pattern of the course of Tb was similar in April, July and October but became more pronounced by the end of the year. In December the body temperature of marmots was rather constant and remained on a lower level, especially at higher ambient temperatures.
2. Metabolic rate during hibernation
To illustrate the energy savings during hibernation we compared the metabolic rate of a single normothermic marmot in winter with hibernating animals. The marmots hibernated at ambient temperatures between -0.5°C and 15°C and showed a minimal and constant metabolic rate between 5°C and 15°C Ta. Below 5°C metabolic rate increased linearly with decreasing Ta. Energy expenditure of hibernating marmots in winter corresponded to about 5.8 % the value the normothermic marmot showed within its winter thermoneutral zone.
DISCUSSION
Alpine marmots exhibit a strong seasonality with regard to thermoregulatory functions like basal metabolic rate, lower critical temperature, extent of thermoneutral zone and body temperature. Considering BMR our metabolic measurements of normothermic marmots throughout the year reveal a seasonal course with highest values found in spring and reduced metabolism in fall and winter. Resting metabolic rate within TNZ well matches values cited in the literature (Armitage et al., 1990) in summer but exceed them in spring. In autumn and winter literature values are higher than those presented in this study. The reduction of energy requirements in fall to 63 % of the spring value in the animal represented supports fattening and is responsible for weight gain prior to hibernation. Measurements of average daily metabolic rate (ADMR) as well as energy intake of marmots show comparable profiles (Körtner, 1991). Energy intake always exceeds energy expenditure and reduction of metabolism in fall allows reduction of energy intake as well, which allows reduced foraging time and energy saving behaviour before entering hibernation. Reduction of thermoregulatory energy demands is non-only achieved by a seasonal rhythm of BMR alone but by a shift of Tlc to lower values and an extension of the thermoneutral zone. Resting metabolic rate at an ambient temperature of 0°C or 10°C - realistic values in an Alpine September is 50 % the value the marmot shows in spring or summer (figure 2). Marmots are less sensitive to cold and living as well as foraging at those temperatures requires half the energy that must be invested early in spring, the time of reproduction and territorial disputes. The shift in lower critical temperature cannot be explained by seasonality of metabolic rate but is due to differences in the regulation of body temperature throughout the year. Marmots increase their body temperature when exposed to ambient temperatures of 30°C except during winter where Tb remained low even during heat exposure. With Ta decreasing below 30°C body temperature as well decreases to a minimum. The extent of the drop in Tb varies throughout the year and is more pronounced in fall than in spring. To give up a constant body temperature regulated within extreme narrow limits is a very energy conserving strategy because metabolic rate can remain minimal unless Tb reaches a lower point and is regulated again and - very important - the gradient between Ta and Tb is reduced. The more pronounced the drop in body temperature the lower is the ambient temperature that induces thermoregulatory response expressed as heat production. As a consequence lowest body temperature mostly coincides with Tlc.
Hibernation plays a dominant role in the marmots strategy for seasonal energy saving but varies in its efficiency with ambient temperature. The lower the temperature in the burrow, the worse the success of hibernation and the greater the weight loss of the marmot (Arnold et al., 1991).
Â
BILANCIO ENERGETICO NELLA MARMOTTA ALPINA DURANTE L'IBERNAZIONE E IN CONDIZIONI DI NORMOTERMIA
Ortmann S. and Heldmaier G.
FB Biologie/zoologie, Philipps-Universitat, D-3550 Marburg
Introduzione
Le marmotte alpine devono far fronte a forti variazioni stagionali della temperatura ambientale e della disponibilità di cibo. Al fine di risparmiare energia, come adattamento al freddo ed alla neve che copre la vegetazione, esse passano i mesi invernali in ibernazione profonda, in gruppi che possono raggiungere 120 individui. Gli animali ibernanti possono immagazzinare riserve di energia, per soddisfare i fabbisogni fisiologici vitali, sia raccogliendo cibo nelle loro tane, sia accumulando grasso corporeo. Le marmotte si affidano unicamente alle riserve di grasso come "carburante" per la stagione dell'ibernazione, che dura circa 6 mesi. I rimanenti sei mesi dell'anno sono dedicati alla riproduzione, ad una intensa e continua assunzione di energia e all'incremento ponderale. Perché quest'ultimo avvenga é necessario che l'energia assunta superi quella consumata. I mammiferi presentano numerosi adattamenti finalizzati a ridurre le spese energetiche, quali, ad esempio, la riduzione della fase di vita attiva e le variazioni del tasso metabolico di acclimatizzazione. Lo scopo di questo studio é di analizzare le variazioni stagionali dei fabbisogni energetici richiesti per la termoregolazione: abbiamo cosi concentrato le nostre attenzioni sulla termo-regolazione e sulle variazioni del tasso metabolico nell'arco di un anno.
Metodi
Le marmotte del nostro laboratorio sono state rinchiuse in un recinto all'aperto, dotato di un'area di riparo. Esse venivano alimentate "ad libitum" con una miscela di granaglie, erba fresca ed acqua. Per la misurazione del lasso metabolico (VO2) gli animali, di volta in volta, venivano trasferiti in una gabbia di plexiglas, con un sottile strato di paglia. In condizioni di normotermia essi venivano dunque sistemati in una stanza climatizzata. Per le marmotte in letargo é stata invece utilizzata una camera climatizzata mobile. I consumi di ossigeno e la produzione di anidride carbonica sono stati misurati in "sistema aperto" (vedi Heldmaier e Steinlechner, 1981); la temperatura corporea (Tb) é stata registrata utilizzano dei termosensori trasmittenti, collegati ad un sistema computerizzato di registrazione.La temperatura ambientale (Ta), misurata con una termocoppia attaccata alla parete della cassa in plexiglas, variava da -0.5°C a +15°C durante l'ibernazione e da -50°C a +30°C durante la normotermia. I valori di VO2, Tb e Ta ad una data temperatura ambientale, venivano misurati ad intervalli di 6 minuti, per almeno 24 ore, durante l'ibernazione; ad intervalli di 1 minuto, per almeno un'ora quando gli animali erano in normotermia. Il VO2 minimo é stato calcolato facendo la media dei tre valori minimi per ogni livello di temperatura. Gli esperimenti in ibernazione sono stati condotti su sette animali durante gli inverni 1988/89 e 1989/90; mentre le misurazioni del metabolismo in normotermia sono state effettuate su quattro marmotte, nei mesi di aprile, luglio, ottobre e dicembre 1990.
Risultati
1. Cambiamenti stagionali di Tb e VO2
La temperatura corporea ed il consumo di ossigeno, a temperatura ambiente compresa tra i -50°C e i +30°C, di una marmotta normotermica, misurati in ottobre, sono illustrati in figura 1. Entrambi i parametri non restano costanti durante l'esposizione al freddo. Il tasso metabolico rimane a valori minimi per valori di Ta compresi tra 0°C e +30°C ed aumenta linearmente con l'ulteriore diminuzione della temperatura ambientale. La temperatura corporea, raggiunto il massimo valore a Ta di +30°C, scende ad un minimo di 35.4°C, per poi aumentare di nuovo, con temperature ambientali inferiori allo 0°C.
In diversi momenti dell'anno sono stati misurati il tasso metabolico a riposo, rapportato al peso corporeo, e la temperatura corporea, a varie temperature ambientali. I risultati ottenuti da un soggetto campione sono rappresentati in figura 2.Nel corso dell'anno si é registrata una caduta dei valori della zona di neutralità termica (TNZ) e variazioni stagionali ritmiche del metabolismo basale (BMR). Quest'ultimo ha fatto rilevare i valori più elevati in primavera-estate ed i più bassi in autunno-inverno. Il peso degli animali varia da 2915 g in primavera a 4220 g in ottobre: non vi é dunque alcuna evidente relazione tra la VO2 peso-specifica e la massa corporea. Durante l'anno, la temperatura minima critica (Tlc) scende da +20°C in primavera a 0°C in autunno, per poi aumentare nuovamente a +20°C in inverno. L'abbassamento del tasso metabolico basale in autunno-inverno riduce i costi energetici della Marmotta alpina, nella zona di termoneutralità , del 63 % rispetto a quelli richiesti in primavera-estate. Come poc'anzi detto, la temperatura corporea non resta costante al variare di quella ambientale. L'andamento della Tb nei mesi di aprile, luglio e ottobre é sovrapponibile; differisce invece verso la fine dell'anno. In dicembre, infatti, la temperatura corporea rimane piuttosto costante e bassa, anche alle temperature ambientali più elevate (Fig. 2).
2. Tassi metabolici durante l'ibernazione
Per evidenziare il risparmio energetico che ha luogo durante l'ibernazione abbiamo confrontato il tasso metabolico di una marmotta normotermica in inverno con quelli di animali ibernanti. Le marmotte sono entrate in letargo a temperature ambientali comprese tra i -0.5°C e +15°C ed hanno evidenziato un tasso metabolico minimo e costante ad una Ta compresa tra i +5°C ed i +15°C. Al di sotto dei +5°C il tasso metabolico aumenta linearmente al decrescere di Ta. Il consumo energetico delle marmotte ibernanti durante l'inverno corrisponde a circa il 5.8 % del valore presentato da una marmotta normotermica, in inverno, nella fase di neutralità termica.
Discussione
Le marmotte alpine presentano forti variazioni stagionali dei meccanismi di termoregolazione, in particolare del tasso metabolico basale, della temperatura critica minima, dell'ampiezza della zona di neutralità termica e della temperatura corporea. Le nostre misurazioni sperimentali, relative all'andamento del BMR, su marmotte normo-termiche, nel corso di un anno, mettono in evidenza variazioni stagionali, con i valori più elevati in primavera ed i valori più bassi in autunno-inverno. I tassi metabolici estivi, a riposo, nella fase di TNZ, da noi rilevati, sono in accordo con quelli citati in letteratura (Armitage et al., 1990); quelli primaverili, al contrario, sono più elevati. I valori autunnali ed invernali citati da altri Autori sono più elevati rispetto a quelli presentati in questo lavoro. La diminuzione dei fabbisogni energetici autunnali, pari al 63 % dei valori primaverili, permette l'accumulo di grassi ed é responsabile dell'incremento ponderale che precede l'ibernazione. I valori del tasso metabolico medio giornaliero (ADMR) e quelli relativi all'energia assunta presentano profili simili (Körtner, 1991). La quantità di energia introdotta supera sempre quella consumata. Il rallentamento del metabolismo in autunno consente alla Marmotta di ridurre la quantità di energia introdotta e, di conseguenza, un risparmio energetico che prelude al letargo. La riduzione dei consumi energetici dovuti alla termoregolazione consegue non solo alla variazione del ritmo stagionale del BMR ma anche all'abbassamento della Tlc e all'ampliamento della zona di neutralità termica. Il tasso metabolico a riposo, a temperature ambientali comprese tra 0°C e +10°C (valori realistici per il mese di settembre, in ambiente alpino), é inferiore del 50 % rispetto a quello relativo al periodo primavera-estate (Fig.2). Le marmotte sono dunque assai poco sensibili al freddo e, d'altra parte, tutte le attività svolte alle basse temperature (tarda estate-inizio autunno), alimentazione compresa, comportano un dispendio energetico che é pari alla metà di quello richiesto all'inizio della primavera, in piena attività riproduttiva, con le relative dispute territoriali. Lo spostamento della temperatura minima critica non può essere spiegato con le variazioni stagionali del tasso metabolico, ma piuttosto con le modificazioni dei meccanismi di termo-regolazione, che si verificano nel corso dell'anno. Le marmotte aumentano la propria temperatura corporea se sottoposte a temperature ambientali di +30°C; questo non si verifica in inverno, quando la Tb rimane bassa anche se le marmotte vengono esposte ad alte temperature. Quando la Ta scende sotto i +30°C anche la temperatura corporea scende al minimo. Il grado di abbassamento della Tb varia nel corso dell'anno ed é più pronunciato in autunno che in primavera. La Marmotta é dunque in grado di modificare i propri meccanismi di termoregolazione, di norma regolati in ambiti molto ristretti. Tale modificazione costituisce un'importante strategia di risparmio energetico, dal momento che i fabbisogni metabolici vengono ridotti al minimo fino a che la Tb non abbia raggiunto i valori critici: solo allora si innesca la termoregolazione. Di fatto, e questo é l'elemento di maggiore importanza, il gradiente tra Ta e Tb viene ridotto. Quanto più é pronunciata la caduta della temperatura corporea, tanto più bassa é la temperatura ambientale che induce una risposta termoregolatrice, in forma di produzione di calore. Ne consegue che la più bassa temperatura corporea pressappoco coincide con la Tlc. L'ibernazione gioca dunque un ruolo fondamentale nelle strategie della Marmotta finalizzate al risparmio energetico stagionale, ma la sua efficacia varia con la temperatura ambientale. Più bassa é la temperatura della tana, più incerto é il successo dell'ibernazione e, di conseguenza, maggiore é la perdita di peso delle marmotte (Arnold et al., 1991).
REFERENCES
ARMITAGE K.B., MELCHER J.C. and WARD J.M. Jr. (1990) Oxygen consumption and body temperature in yellow-bellied marmot populations from montane-mesic and lowland-xeric environments. J. Comp. Physiol. B, 160: 491-502.
ARNOLD W., HELDMAIER G., ORTMANN S., POHL H., RUF T. and STEINLECHNER S. (1991) Ambient temperatures in hibemacula and their energetic consequences for Alpine marmots (Marmota marmota). J. therm. Biol., 16,4: 223-226.
HELDMAIER G. and STEINLECHNER S. (1981) Seasonal central of energy requirements for thermoregulation in the Djungarian hamster (Phodupus dsungorus), living in natural photoperiod. J. Comp. Physiol. B, 142: 429-437.
KORTNER G. (1991) Winterschlaf und Seine Auswirkungen auf den Energiehaushalt beim Alpenmurmeltier (Marmota marmota). Dissertation, Philipps-Universitat, Marburg/Lahn.
Fig. 1: Metabolic rate (above) and body temperature (below) of a single normothermic marmot exposed to a stepwise decrease in ambient temperature. The lines connect values of resting metabolic rate or constant Tb at a given Ta.
ritorno/back
Fig. 2: Metabolic rate (above) and body temperature (below) of a normothermic marmot tested at four different times the year.
ritorno/back
This study was supported by the Deutsche Forschungsgemeinschaft (SFB 305)
ritorno/back
Tornare index / back contents
Â