Fig. 1:View of the area of study, reproducted from a photograph.

THE MARMOT POPULATION OF THE TUSCAN-EMILIAN APENNINE RIDGE
Sala I., Sola C., Spampanato A. and Tongiorgi P.
Dipartimento di Biologia Animale, Università di Modena, via Universita 4, 1-41100 Modena.
Abstract - In the Modena Apennines above the tree line (1600 -1700 metres a.s.l.), the marmot is today found almost everywhere. The present population is the result of introduction from 1946 onwards. The Spread of this rodent in an environment different to its original alpine habitat offers numerous points of scientific and administrative interest and justifies the eco-ethological study of this population. Since 1988, the daily and seasonal activity rhythms and the social structure of a group of marmots on the northern slopes of the Monte Cimone have been observed. (This study was funded by the Province of Modena, Department of Agriculture, Fish and Hunting and by the Italian Ministero dell'Università e della Ricerca Scientifica e Tecnologica, MURST 60% programme).
Since the 1950's, the marmot (Marmota marmota L.) has been introduced on a number of occasions in various regions of the northern Apennines and is now spread, although not continuously, in the mountainous area from Parma to Bologna (Ferri et al., 1988).
In1987, marmot settlements along the Monte Cimone-Libro Aperto (Modena) ridge were trapped in detail and the location of burrows described in terms of the major environmental characteristics (altitude, exposure, geomorphology etc.). Irrespective of the exposure of the slope, the majority of the area above the line-tree was found to be colonized. Flatter and less steep areas where the substrate is composed primarily of earthy deposits are only rarely colonized, while very rocky parts are not inhabited. Settlement is also affected by vegetation; the extensive vaccinium (bilberry, Vaccinium myrtillus), for example, is carefully avoided (Ferri et al., 1988).
It is known that environmental conditions (altitude, size of territory, availability of food) affect reproduction and, consequently, also the Social structure of the populations (cf. Barash, 1973, 1974a, b). From this point of view, the recently established Apennine marmot population which has settled in an environment somewhat diverse to its original one is of interest for a number of reasons.
Below, we report the results of a three-year study of the density, population structure and dynamics, daily and seasonal activity rhythms, behaviour and the utilization of territory of a marmots group living on the Apennine.
STUDY AREA AND METHODS
The study was carried out on a number of marmot families settled on the northern slopes of Monte Cimone in an area of about 5 hectares at an altitude between 1800 and 1950 metres. The area is characterized by grassy banks and slopes interspersed with rocky outbreaks and gorges, delimited above by a series of rocky crags and below by the flat grassland of the Pian Cavallaro. The vegetation is typical of the Apennine grazing land above the line-tree (1600 - 1700 metres a.s.l.), where predominantly gramineae areas alternate with the vaccinium. Burrows were marked with numbered metal pegs (Fig. 1) such that they could be localized from a distance and in following years. Every year from 1988 onwards, the animals in the area of study were caught with snares and individually marked. The weight and sex of each animal caught was recorded. Four age groups were distinguished: 1) young, 2) yearlings, 3) two-year olds, 4) adults. Animals in groups 3 and 4 could only be distinguished on the basis of data and markings from previous years.
In order to identify the marmots from a distance, coloured plastic ear clips (Dalton mod. Minirototag 20x5 mm) were hung to the animals' ears and their fur was tinted with hair colouring. Ink tattoos made with ACS 5 pincers and cold marking with freezing gas spray (-40° C) were used for long-term individual recognition. Although sometimes difficult to read, the coloured ear clips proved relatively efficient, but rarely survived for longer than a season. Cold marking had no effect on adults. Results were better with young, but still did not allow easy recognition of the animals. Observations were carried out from May to the end of September in alternate weeks for 2-4 consecutive days, beginning at down before the animals leave the burrow and continuing until all return in the evening. The position and activity of the individual animals (Scanning Sample Method, Altman, 1974), atmospheric conditions and air temperature were recorded every 15 minutes.
A total of 607 hours of observation were carried out on 57 days.
The following codes were used to describe the activity of the marmots: 1. X - Animal in burrow; 2. D - Animal in movement; 3. S - Animal sedentary ; 4. B - Sunni ng; 5. A - Alert; 6. C - Up-alert; 7. M Eating; 8. F - Calling out; 9. L - Fighting; 10. G - Playing; 11. E - Escape; 12. I - Friendly interaction; 13. P - Grooming; 14. Q - Auto-grooming ; 15 . R - Chasing (not in play); 16 . T - Carrying grass; 17. N - Hidden individual movement; l8. Z - Burrowing; 19. O - Marking with scent; 20. U - Recumbent test; 21. K - About to enter burrow. In order to simplify the analysis of the frequency of the individual activities, the above 21 behavioural categories were regrouped into the following 6 categories. The figures in brackets indicate the classifications contained in each category:
I - Feeding (7)
II - Rest (3, 4, 20)
III - Movement (2, 11, 16, 18, 21)
IV - Interaction (10, 12, 13)
V - Alert (5, 6, 8)
VI - Other (14, 17, 19)
In the statistical results, data for one and two-year olds has been combined under " yearlings".
Tab. 1: Prospects of the specimens resident in the study area from 1988 to 1991
RESULTS AND DISCUSSION
In the first year (1988), six animas, all over two years of age, were caught (identified as A-F). In May 1989, approximately fifteen days after the end of hibernation, five marmots were seen at the same time outside. In June, two couples remained, each occupying its own family territory. Both the male (indicated as 02) and the female (03) of one couple were caught and marked, while of the other couple (female A and male B of 1988), only the female was captured. In July, five offspring of the couple A-B appeared, four of which, three females and one male, were caught and marked (05, 06, 07, 08). On 15 May 1990, five young of the previous year were observed in the study area, but it was non possible to capture them. Five adult marmots, three of which were re-caught and marked (A, 02, and 03), were also seen. In the same period and following wild combat, the male 02 was expelled from its territory by an intruder identified as Z. The couple A-B reproduced five further offspring (four females and one male) who were all caught and marked (10, 11, 12, 13, 14).
In May 1991, the couple 03-Z still occupied its territory, while in the territory of A-B only the five animals born in 1989 were seen. The couple 03-Z reproduced four offspring. The population in the area recorded in the summer thus increased from 7 animals in 1988 to 9 in 1989 and 14 in 1990, but fell to 11 in 1991. The particular demographic situation revealed in 1991 appears to confirm that also in the Apennines climatic conditions immediately following the end of hibernation play an important role in population regulation. Indeed in 1991, heavy snowfall directly after winter hibernation around 20 April left a complete covering of snow which persisted for more than a month and restricted the feeding land of the marmots which were already active.
In the area of study and in those adjacent of the Monte Cimone, the marmots occupy clearly defined family territories which they actively defend against intruders. Only occasionally do the yearlings and young of one family enter neighbouring territory, and only then the outer edges, and do not experience particular aggression on the part of the residents. It can thus be said that the families in the area of study on the Monte Cimone do not constitute a colony (cf. Mann & Janeau, 1988). The size of the family territories on the Monte Cimone (22.5 hectares) is comparable to those of the Alpine populations: 1.4-2 hectares according to Zelenka (1965); 2-5.7 hectares for Mann & Janeau (1988); on average 3.6 hectares in the Gran Paradiso park according to Lenti (1988). Habitation in different burrows in the summer and the winter, which has been frequently seen in the Alps (Couturier, 1964; Zelenka, 1965; Barash, 1976; Huber, 1978; Durio, 1988; Lenti, 1988), does not seem to be the case in the Apennines. Of the four families observed over the last years in the area of study and surrounding territories, two used different burrows, while the other two remained in the same burrow throughout the year for both reproduction and hibernation. Behavioural observations were frequently disturbed by adverse meteorological conditions which lead to difficulties both in on site observation and in the subsequent statistical analysis. Nevertheless, results over the years remained for the main part consistent.
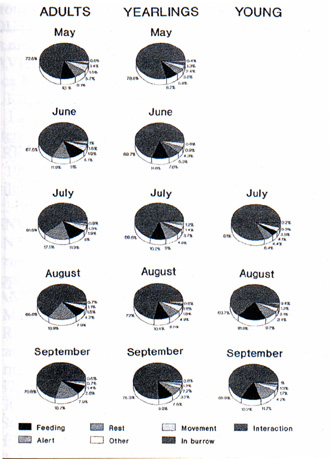
Marmots in all age groups (adults, one and two-year olds and young) spend the majority of their time in the burrow (Fig. 2). In general, adults spend more time outside than the yearlings, and these more than the young.
In adults, time spent outside is primarily dedicated to test (14.5%). 9.7% of their time is spent feeding, 4.5% in movement and 3.6% in other activities (social interaction, surveillance, marking of territory etc.). In the yearlings and young, on the other hand, the main activity is feeding (12.4% and 8.2%, respectively). The yearlings spend the same amount of time in social interaction as in movement across the territory (3.8%) (Fig. 2). Feeding increases progressively up to the end of July in adults and up to the end of August in others, and then falls in subsequent months (Fig. 3). Friendly interaction (play, grooming etc.) is particularly intense among young, especially in July immediately following emergence from the natal burrow. For the main part, this interaction is amongst themselves or with the mother, rarely with the father.
Following emergence of the young, increased vigilance on the part of the parents was noted. The incidence of friendly interaction (play) is still high amongst the yearlings of the same age. Aggressive interaction was seen primarily in spring and only amongst adults of different families. On average, marmots become active about half an hour after dawn. Rainfall affects the onset of morning activities, whereas fog and wind show no significant influence. Return in the evening is determined to a greater extent by the weather. In general, adults are outside in the early part of the morning and in the late afternoon, while the activity of the yearlings and even more so of the young is more evenly distributed across the whole day (Fig. 4). In agreement with the findings of Arnold (1988) in M. marmota and Armitage (1976) in M. flaviventris, the observations suggest that in the Apennine population there is also a correlation between daily activity rhythms, temperature of the environment and size of the animal. This first report on marmot behaviour complements recent observations of the alpine populations carried out by Swiss, French and German researchers (cf. Arnold, 1988a, b, 1990a, b; Mann & Janeau 1988; Solari 1988). The aims of future observations are: 1) an evaluation of family group dynamics and the establishment on the basis of an analysis of individual histories of the reproductive strategies of this large rodent; 2) determination of the use that the animal makes of its territory and a better understanding of the existing relationships between behaviour and environmental variables.
INDAGINI SUL POPOLAMENTO DI MARMOTTA DEL CRINALE APPENNINICO TOSCO-EMILIANO
Sala I., Sola C., Spampanato A. and Tongiorgi P.
Dipartimento di Biologia Animale, Università di Modena, via Universita 4, 1-41100 Modena.
A partire dagli anni 1950, la marmotta (Marmota marmota L.) è stata introdotta più riprese in varie località dell'Appennino settentrionale ed è oggi diffusa, seppure in modo discontinuo, dal parmense al bolognese (Ferri et al.,1988).
Nel 1987 gli insediamenti di marmotte localizzati lungo il crinale monte Cimone-Libro Aperto (Modena) sono stati mappati dettagliatamente e le aperture di tana sono state censite e caratterizzate in rapporto ai principali parametri ambientali (altitudine, esposizione, geomorfologia ecc.). Gran parte della fascia altitudinale ai di sopra del limite della vegetazione arborea è risultata colonizzata, indipendentemente dall'esposizione dei versanti. Le aree a scarsa pendenza e quelle pianeggianti, dove il substrato è essenzialmente costituito da depositi terrosi, sono colonizzate solo raramente, mentre sono evitate quelle dove affiorano estesi banchi rocciosi. Anche le associazioni vegetali condizionano gli insediamenti: il vaccinieto (Vaccinium myrtillus) esteso è, ad esempio, accuratamente evitato (Ferri et al., 1988). Come noto, le condizioni ambientali (altitudine, ampiezza del territorio, disponibilità di risorse alimentari ecc.) influiscono sulle strategie riproduttive e quindi sulla struttura sociale delle popolazioni (cfr. Barash, 1973, 1974a, b). La popolazione di marmotte dell'Appennino, di recente costituzione e insediata in un ambiente relativamente diverso da quello originario, offre, sotto questo aspetto, numerosi motivi di interesse. Vengono qui riferiti in sintesi i risultati di una indagine triennale tesa ad acquisire informazioni circa densità, struttura e dinamica di popolazione, ritmi di attività circadiana é stagionale, utilizzazione del territorio e alcuni aspetti del comportamento di un nucleo di marmotte dell'Appennino.
Area di studio e metodi
Lo studio ha interessato alcune famiglie di marmotte insediate sui versante nord del Monte Cimone, in un'area di circa 5 ettari tra 1800 e 1950 m di quota, costituita da una serie di dossi e pendii erbosi intervallati da canaloni e affioramenti rocciosi. L'area di studio è delimitata superiormente da una serie di balzi rocciosi mentre in basso continua con le praterie pianeggianti di Pian Cavallaro. La vegetazione è quella tipica dei pascoli appenninici oltre il limite della vegetazione arborea (1600-1700 m slm), dove aree dominate da graminacce si alternano al vaccinieto. Le aperture di tana presenti nell'area di studio sono state contrassegnate con picchetti metallici numerati Fig. 1) in modo da permetterne la lettura a distanza e in anni successivi.
A partire dal 1988, ogni anno, gli individui residenti nell'area sono stati catturati per mezzo di lacci e marcati per permettere il riconoscimento individuale. Di ogni esemplare catturato sono stati rilevati peso e sesso. L'età veniva attribuita distinguendo quattro categorie: 1) nati nel'anno in corso; 2) giovani dell'anno precedente; 3) individui nel secondo anno di vita e 4) adulti.
L'attribuzione degli animali alla terza o alla quarta categoria è stata possibile solo in base ai dati di cattura e marcatura degli anni precedenti. Per l'identificazione a distanza, le marmotte sono state marcate con clips auricolari di plastica colorata (Dalton mod. Minirototag, 20x5 mm) e mediante coloritura del pelo con tinture per capelli. Per il riconoscimento individuale a lungo termine sono stati effettuati tatuaggi ad inchiostro con pinzeACS 5 e marcatura a freddo con gas refrigerante spray (-40° C). Le clips auricolari colorate si sono rivelate abbastanza valide, anche se tal volta di difficile lettura, ma difficilmente permangono per più di una stagione di attività. La marcatura a freddo non ha avuto alcun effetto sugli adulti, mentre sui piccoli dell'anno ha dato risultati migliori, ma che non permettono comunque un agevole riconoscimento degli animali. Le osservazioni sono state condotte da maggio alla fine di settembre, a settimane alterne, per 2-4 giorni consecutivi, dall'alba, prima dell'uscita degli animali dalle tane, fino al rientro definitivo di tutte le marmotte per if riposo notturno. Ogni 15 minuti veniva registrata posizione e attività dei singoli individui (Scanning Sample Method, Altman,1974), le condizioni atmosferiche e la temperatura dell'aria. Complessivamente, sono state eseguite 607 ore di osservazioni in 57 giorni.
Le attività svolte dalle marmotte sono state codificate secondo le seguenti categorie: I. X - animale non in superfice; 2. D - animale in movimento; 3. S - animale seduto; 4. B - bagno di sole; 5. A - alterna; 6. C posizione a candela; 7. M - mangiare; 8. F - animale che emette grida; 9. L - lotte di tipo conflittuale; 10. G - gioco; 11. E fuga; 12. I - interazioni amichevoli; 13. P - grooming; 14. Q auto-grooming; 15. R - rincorrersi (non per gioco); 16. T - trasporto erba; 17. N - individuo momentaneamente nascosto; 18. Z scavare; 19. O - marcare con stimoli odorosi; 20. U - posizione di riposo steso; 21. K - individuo in atto di scomparire in una tana. Per semplificare l'analisi delle frequenze delle singole attività, le 21 attività precedentemente indicate sono state raggruppate in sei sole categorie.
Per ciascuna di esse è indicato, fra parentesi, il codice delle attività in esse compreso.
I - Alimentazione (7)
II - Riposo (3, 4, 20)
III - Movimento (2, 11, 16, 18, 21)
IV - Interazioni (10, 12, 13)
V-Allarme (5, 6, 8)
VI - Altre (14, 17, 19)
Nell'elaborazione statistica, i dati relativi agli individui di 2 e 3 anni di età sono stati cumulati sotto la voce "giovani" (Tab. 1).
Risultati e discussione
Nel primo anno (1988) sono stati catturati sei individui (ind. A-F), tutti di età superiore ai due anni (adulti e subadulti) (Tab. 1).
Nel maggio 1989, circa quindici giorni dopo l'uscita dal letargo, sono state osservate cinque marmotte contemporaneamente in superficie in giugno rimanevano solo due coppie, ognuna delle quali dislocata in un proprio territorio familiare. Di una coppia sono stati catturati e marcati ambedue gli individui, indicati come 02 (maschio) e 03 (femmina), mentre dell'altra (femmina A e maschio B del 1988) è stata catturata solo la femmina. ln luglio, sono comparsi in superficie cinque piccoli nati dalla coppia A-B, quattro dei quali, tre femmine ed un maschio, sono stati catturati e marcati (indiv. 05, 06, 07, 08).
If 15 maggio 1990, nell'area di studio sono state osservati i cinque giovani nati nell'anno precedente, che non è stato possibile ricatturare, e cinque marmotte adulte, di cui solo tre sono state ricatturate e marcate (A, 02 e 03). Nello stesso periodo, dopo un cruento combattimento, il maschio 02 è stato espulso dal suo territorio da un estraneo denominato Z. La coppia A-B si é nuovamente riprodotta, dando alla luce cinque piccoli (quattro femmine e un maschio) che sono stati catturati e marcati (ind. 10, 11, 12, 13, 14).
Nel maggio 1991 la coppia 03-Z occupava ancora il proprio territorio, mentre nel territorio della coppia A-B sono stati osservati esclusivamente i cinque individui nati nel 1989. La coppia 03-Z si è poi riprodotta generando 4 piccoli.
La popolazione dell'area di studio, rilevata nel periodo estivo, è passata quindi dai 7 individui del 1988, ai 9 del 1989, ai 14 del 1990, per calare poi a 11 nel 1991. La particolare situazione demografica riscontrata nel 1991 sembra confermare, anche sull'Appennino, l'importanza delle condizioni climatiche nel periodo immediatamente successivo al risveglio dal letargo nella regolazione delle popolazioni di marmotta. Infatti, nel 1991, subito dopo l'uscita dal letargo invernale, registratasi intorno al 20 aprile, abbondanti nevicate hanno determinato una copertura nevosa continua, che si è protratta per oltre un mese e che ha impedito il pascolo alle marmotte già attive.
Sul Monte Cimone, nell'area di studio e nei territori adiacenti, le marmotte costituiscono nuclei familiari occupanti territori nettamente distinti tra loro e attivamente difesi dall'ingresso di estranei. Solo occasionalmente giovani o piccoli di una famiglia penetrano nei territori confinanti, ma solo nelle zone marginali, senza suscitare particolare aggressione da parte dei residenti. É possibile dunque affermare che i nuclei familiari presenti nell'area di studio del Monte Cimone non costituiscono una "colonia" (cfr. Mann & Janeau, 1988).
L'estensione dei territori familiari nell'area di studio del Monte Cimone, 2-2, 5 ettari, è comparabile a quella delle popolazioni delle Alpi: 1,4-2 ettari secondo Zelenka (1965); 2-5, 7 ettari secondo Mann e Janeau (1988) i mediamente 3, 6 ettari nel Parco del Gran Paradiso secondo Lenti (1988).
La frequentazione di tane distinte in inverno e in estate, rilevata frequentemente sulle Alpi (Couturier, 1964; Zelenka, 1965; Barash, 1976; Huber, 1978; Durio, 1988; Lenti, 1988), non sembra costituire la regola sull'Appennino: infatti, delle quattro famiglie osservate nel corso degli ultimi anni nell'area di studio e nei territori contigui, due coppie hanno utilizzato tane diverse, mentre te altre due hanno frequentato tutto l'anno la stessa tana, sia per il parto sia per l'ibernazione. Le osservazioni etologiche sono state spesso disturbate dalle avverse condizioni metereologiche: cio ha comportato, da un lato, difficoltà sia in sede di osservazioni di campagna, sia nella successiva fase di elaborazione dati. Nonostante cio, è emersa una sostanziale concordanza nei risultati nel corso degli anni.
Le marmotte di tutte e tre le classi d'età (adulti, giovani di 2-3 anni e nati nell'anno) trascorrono la maggior parte del giorno all'interno della tana (Fig. 2). In generale, gli adulti trascorrono più tempo in superficie dei giovani e questi, a loro volta, dei piccoli. Il tempo trascorso dalle marmotte adulte fuori della tana è prevalentemente dedicato al riposo (14,5%); all'alimentazione dedicano il 9,7% del tempo, il 4,5% è speso per gli spostamenti, mentre if 3,6% è impiegato in altre attività (interazioni sociali, sorveglianza e marcatura del territorio ecc.).
Nei giovani e nei piccoli, invece, l'attività prevalente è l'alimentazione (rispettivamente 12,4% e 8,2%); nei giovani il tempo dedicato alle interazioni sociali eguaglia quello dedicato agli spostamenti attraverso il territorio (3,8%) (Fig. 2).
L'attività alimentare aumenta progressivamente fino a luglio negli adulti e fino ad agosto negli altri, per decrescere nei mesi successivi (Fig. 3). Nei piccolilte interazioni amichevoli (gioco, grooming ecc.) risultano particolarmente intense, specialmente in luglio subito dopo l'uscita dalla "tana parto", principalmente tra loro e con la madre, mentre le interazioni col padre sono scarse. In corrispondenza delle prime uscite dei piccoli, é stato osservato negli adulti un incremento dell'attività di vigilanza. La frequenza di interazioni amichevoli (gioco) è ancor più elevata nei giovani coetanei. Interazioni aggressive sono state osservate principatmente in primavera, ma solo tra adulti di famiglie diverse. Le marmotte iniziano la loro attività mediamente mezz'ora dopa l'alba. L'inizio dell'attività mattutina è condizionato da eventuati precipitazioni; nebbia e vento non influiscono significativamente. Il rientroserale è invece maggiormente influenzato dalle condizioni del tempo. In generale, gli adulti sono presenti in superficie soprattutto nella prima parte della mattina e nelle tarde ore pomeridiane, mentre l'attività dei giovani e, ancor più, quella dei piccoti è più uniformemente distribuita nell'arco della giornata (Fig. 4). Le osservazioni sui ritmi giornalieri di attività suggeriscono anche per la populazione appenninica, una correlazione tra ritmo di attività giornaliera, temperatura ambientate e taglia dell'individuo, analogamente a quanto registrato da Arnold (1988) in M. marmota e da Armitage (1976) in M. flaviventris. Questo primo contributo alla determinazione del profito comportamentale della marmotta integra precedenti osservazioni effettuate in questi ultimi anni su popolazioni dell'arco alpino da ricercatori svizzeri, francesi e tedeschi (cfr. Arnold, 1988a, b, 1990a, b; Mann & Janeau 1988; Solari 1988). Obiettivo delle osservazioni negli anni futuri sono: la valutazione della dinamica dei gruppi familiari e l'accertamento, attraverso l'analisi delle storie individuali, delle strategie riproduttive esibite da questo grosso roditore; stabilire quale uso egli faccia del territorio e pervenire ad una migliore comprensione delle relazioni esistenti tra manifestazioni comportamentali e variabili ambientati.
Fig. 1:View of the area of study, reproducted from a photograph.
Fig. 3: Mean frequency of the time spent in the different activities by the three age groups.
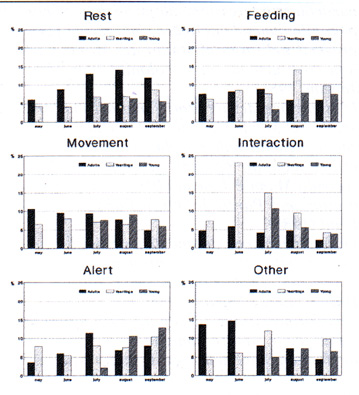
Fig. 4: Daily actvitity cycle of young, yearling and adult marmots.
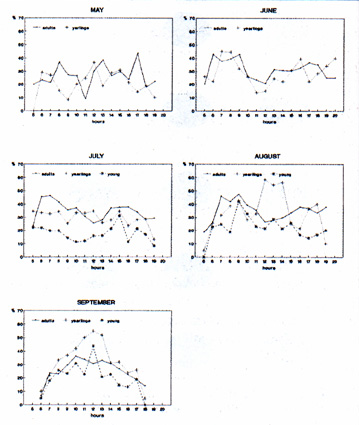
Table 1. Prospects of the specimens resident in the study area from 1988 to 1991.
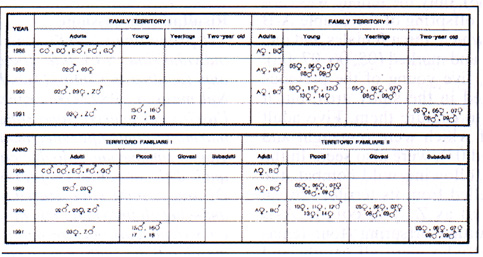
| YEAR | Family territory I : Territorio familiare I | Family territory II : Territorio familiare II | ||||||
| Adults/Adulti | Young/Piccoli | Yearling/Giovanni | Two-year old/Subadulti | Adults/Adulti | Young/Piccoli | Yearling/Giovanni | Two-year old/Subadulti | |
| 1988 | Om, Dm, Em, Fm, Gm | Af, Bm | ||||||
| 1989 | 02m, 03f | Af, Bm | 05f, 06f, 07f, 08m, 09m | |||||
| 1990 | 02m, 03f, Zm | Af, Bm | 10f, 11f, 12m, 13f, 14 f | 05f, 06f, 07f, 08m, 09m | ||||
| 1991 | 03f, Zm | 15m, 16m, 17, 18 | 05f, 06f, 07f, 08m, 09m | |||||
REFERENCES
ALTMANN J., 1973 - Observational study of behavior: sampling methods. Behaviour, 49: 227-267.
ARMITAGE K. B., DOWNHOWER J. F., SVENDSEN G. E. 1976 - Seasonal changes in weights of marmots. Ann. Midi. Nat., 96: 36-51.
ARMITAGE K. B., 1987 - Social dynamics of mammals: reproductive success, kinship and individual fitness. Trends Ecol. Evol., 2: 279-283.
ARNOLD W., 1988b - Thermoregulation as a limit to habitat use in alpine marmots (Marmota marmota).
0ecologia, 76: 544-548.
ARNOLD W., 1990a - The evolution of marmots sociality: why disperse late? Behav. Ecol. Sociob., 27: 229-237.
ARNOLD W., 1990b - The evolution of marmots sociality: losts and benefits of joint hibernation. Behav. Ecol. Sociobiol., 27: 239-246.
BARASH D. P., 1976 - Social behaviour and individual differences in free living alpine marmots (Marmota marmota). Anim. Behav., 24: 27-35.
BARASH D. P., 1989 - Marmots, social behavior and ecology. Stanford University Press, 360 pp.
BOPP P., 1955 - Kolonialterritorien bel Murmeltieren. Rev. suisse Zool., 62: 295-299.
BOPP P., 1956 - Zur Topographie eines Kolonialterritoriums bei Murmeltieren. Rev. suisse Zool., 63: 255-26.
CAGNOLARO L., SPAGNESI M, 1981 - La Marmotta (Marmota marmota marmota L. 1758). In:
Distribuzione e biologia di 22 specie di mammmiferi in Italia. Roma, C.N.R- Collana Progetto Finalizzato "Promozione della Qualita dell'Ambiente" AQ/1/142164, pp. 31-37.
COUTURIER M-A-J., 1964 - La Marmotte des Alpes (Marmota marmota marmota (L.) 1758). In: Le gibier des montagnes françaises, (10 ediz., 1981). Grenoble, Arthaud, pp. 137-189.
DAVIS D. E., 1982 - Woodchuks. In: Handbook of census methods for terrestrial vertebrates. CRC Press, Boca Raton; Florida, pp. 147-147.
DURIO P., BASSANO B., PERRONE A. 1987 - Le tane della marmotta (Marmota marmota L.). In: Incontro su: La marmotta alpina (Marmota marmota). Ente Parco Nazionale Gran Paradiso, Gran Nomenon, 24-25 settembre 1987, pp. 25-31.
FERRI M., PIGOZZI G., SALA L., SOLA C., SPAMPANATO A., TARANTINO J., TONGIORGI P., TOSI L., 1988 - Primi risultati di una ricerca sulla popolazione di Marmota marmota del crinale appenninico tosco-emiliano. Atti "I Convegno Nazionale dei Biologi della Selvaggina". Bologna 28-30 gennaio,1988. Suppl. Ric. Biol. Selvaggina, 14: 339-349.
KENWARD R., 1987 - Wildlife radio tagging. Academic Press, London.
LENTI D., 1988 - Cattura, marcatura e immobilizzazione farmacologica della marmotta alpina (Marmota marmota L.). Atti "I Convegno Nazionale dei Biologi della Selvaggina". Bologna 28-30 gennaio, 1988. Suppl. Ric. Biol. Selvaggina, 14: 353-364.
MANN C. S., JANEAU G., 1988 - Occupation de l'espace, structure sociale, et dynamique d'une population de Marmottes des Alpes (Marmota marmota L.). Gibier Faune Sauvage, 5 : 427-445.
PERACINO V., BASSANO B., 1987 - La Marmotta (Marmota marmota L.) nel Parco Nazionale del Gran Paradiso: metodologie di censimento e valutazione della sua distribuzione (1986 -1987). In: Incontro su: La mamotta alpina (Marmota marmota), Ente Parco Nazionale Gran Paradiso, Gran Nomenon, 24-25 settembre 1987, pp. 13-24.
PIGOZZI G., 1984 - The den system of the Alpine Marmot (Marmota marmota marmota) in the National Park of Stelvio, Northern Italy. Z. Saugetierk., 49: 15-21.
PREMUDA G., 1975 - Le marmotte del Lago Nero. Diana, 11 : 28-31.
SOLARI C., 1988 - Distribution and population density of the Alpine marmot (Marmota m. marmota) in the Canton of Ticino (Switzerland). A causal analysis. J. Jagdwiss., 34: 77-85.
ZELENKA G., 1965 - Observations sur l'ecologie de la marmotte des Alpes. Terre et Vie, 19: 238-256.
ritorno/back