ANALYSIS OF THE TEMPORAL AND SPECTRAL ACOUSTIC PARAMETERS OF Marmota marmota SOUND EMISSIONS
Bassano°, Durio P.°, Facette C.*, Macchi E.°, Piazza R.*, Trinchero C.*
° Dipartimento di Produzioni Animali, Epidemiologia ed Ecologia - Università di Torino. Via Nizza 52, 10126 Torino, Italy
* Dipartimento di Morfofisiologia Veterinaria - Università di Torino. Via Campana 16, 10126 Torino, ltaly
Abstract - Vocalisations of marmots (Marmota marmota) of different colonies of two valleys of the Western Alps have been analyzed. The vocal emissions were recorded lying in wait, under natural conditions, and also after artificial stimulations (realising moulds of birds, volontary presence of humans carrying lights and providing the capture of two animals). The physical analysis of the vocalisations permitted to releave at least 4 different types of emissions: Single Whistles and Multiple Whistles, Single and Multiple Squeaks, Growls and Screams (emissed during capture). The emissions were analyzed in their temporal and spectral characters, and from the analysis it results that whilst the whistles and squeaks have harmonic structure with discrete spectral components added in components of low or null noise level, the screams and growls are polluted by noise, produced by the subject, in high level besides which consents the finding of the harmonic structure. Such a difference is analysed from a semantic point of view.
l INTRODUCTION
From the literature concerning sound emissions of marmots (Lenti Boero D., 1988-89), in particular the alarm calls, it results that, from the point of view of the motivation, they must be studied on the ground of various interpretative models, amongst which we remember those that are based on the following hypotheses:
a) the semantics hypothesis, that implicates the fact that in the animal there exists a form of representation of the external world, from which one or more signals arrive to which follows in a temporal manner a behavioural answer;
b) the excitement hypothesis for which in the signal emitted by animal is codified a particular emotional condition "determinated" by the external context;
c) the fraud hypothesis with two possibilities for the animal: 1°) signal an inexistent predator in order to take advantage of the general flight for the possessment of unguarded resources; 2°) not signal an existent predator permitting it to act undisturbed. In the paper (Noyes D.H., Holmes W.G., 1979) the authors describe the observation of the different behaviour of the marmots in relation to the greater or minor of risk gravity. The behavioural stimulus was constituted by a radio-controlled ersatz golden Eagle. It is evident that the answer to the question about the motivation, in various contextual situations, requests a detailed study of the physical characteristics of the emission, repeated for each type of emission, so as the analysis of parameters in relation to the contextual data may give an answer in terms of a multivaried statistic analysis, on the existence or non-existence of a relation cause/effect between the external signal (or the group of external signals) and the behaviour of subject, defined by the whole values of a certain number of acoustic parameters of the signal. In this sense, the present research wills to provide a contribution about emissions of Marmota marmota. In (Lenti Boero D., 1988-89) we find on Marmota marmota the description of the following types of alarm calls: Multiple whistle (MW), definite as "shrill sound consisting in a succession of elementary signals at high frequency emitted in a succession more or less rapid". Noticed when terrestrial animals or men approach.
Single whistle (SW), unique signal that "isn't repeated", emitted on the occasion of the aerial passage of potential predator;
Multiple slackened whistle (MSW), defined as a signal "more similar to a squeak than to a real whistle "of a more low tonality, of a not yet clarified motivation; in the present report it will be refereed to as "squeak".
Yelp, sound "similar to the yelping of a dog";
Growl, sound "similar to the growling of a dog";
Tooth chatter, (Waring G., 1966) "a rapid, non-vocal, clicking sound resulted by antero posterior and viceversa movements of the lower jaw, screams, (Waring G., 1966) "emissions obtained by prolonging single whistle".
In the present report we have called "screams", particular voiced signals emitted during the capture.
In the present research have been analysed single and multiple whistles, squeaks, growls, screams besides the composition of the signals emitted during and after the capture.
It is recalled that, by a phonation point of view, whilst for the single and multiple whistle (and probably for the squeaks) the vocal chords aren't in vibration, for the other signals there is vibration. We arrive at this conclusion on the basis of the data relieved of the fundamental frequency of the voiced signals through comparison between the dimensions of the humans' and of the marmots' vocal tracts, dimensions that can be considered about inversely proportional to the medium fundamental frequency.
2- METHODS AND MATERIALS
The vocalisations of Marmota marmota have been recorded with the use of a Uher 4400 tape recorder provided with unidirectional microphone and parabola.
The recordings have been affected in two colonies posted at different altitudes (1700 and 1740 m above sea-level) inside and at the borders of the Parco Naturale Orsiera Rocciavre (Western Alps) and in a colony present in the Vallone of the Ischiator (1791 m above sea-level), Valle Stura (Province of Cunco-Western Alps).
In the first two zones of study (Parco Orsiera), in order to incite the vocal reaction of the marmots, moulds of differing species of birds have been utilized, predators and not (royal Eagle - Aquila chrysaetos-, buzzard - Buteo buteo-, imperial Crow - Corvus corax-, Falconidae, alpine chough - Pyrrhocorax graculus-, Jay - Garrulus glaudarius-) that we made to fall along a cord 20 meters long overhung above the marmots' burrows.
To make these operations possible two colonies were chosen based on rocky surfaces, from which from the top the mould were released. The length of the suspend cord was respectively of 180 and 250 m.
The starting point of the ersatz must be hidden to the view of the marmots. In fact the animals that saw the operator release the ersatz never emitted acoustic signals. Voiced emissions were also emitted during the sudden appearance of fog.
In the 3rd area of study (Stura Valley), we have recorded vocalisations of animals captured with traps and manipulated. Voiced emissions provoked by the passage of dogs and people, that however resulted not technically utilizable. The physical analysis of the emissions have been effectuated with a Digital sonagraph 7800 connected with interface 7812-488 to a compatible IBM PC provided with programs for supplying time wave with different scales, sonograms of the signals with maximum frequency either 6400 or also 12800 Hz and spectra either with temporal regulable intervals from 5 to 160 ms, and also globals for prefixed intervals up to 2500 ms.
3- EXPERIMENTAL DATA ANALYSIS
As already mentioned, the research has regarded whistles (single and multiple), squeaks (single and multiple), growls and screams. Since the last two signals have been relieved during the capture we have emissions of screams alternated to growls, the two signals will be treated together.
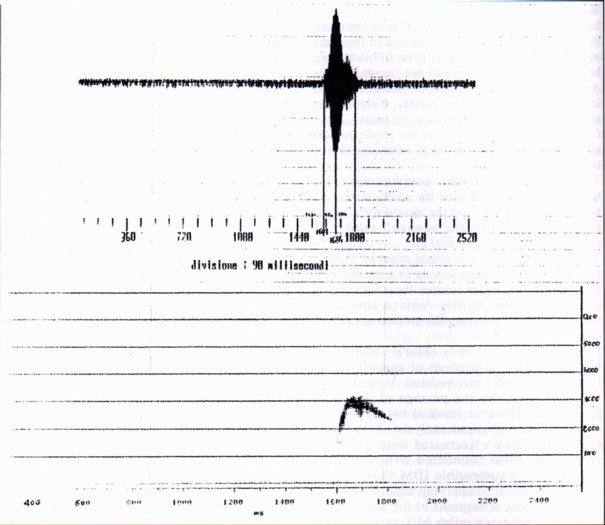
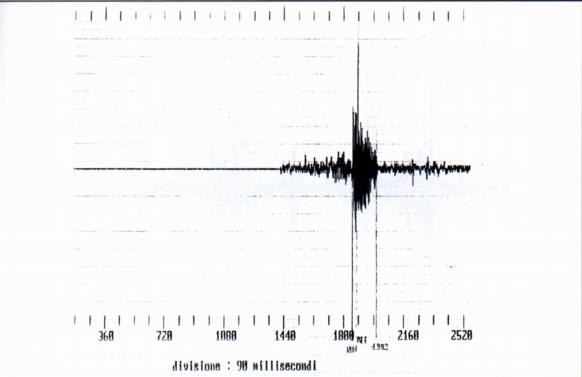
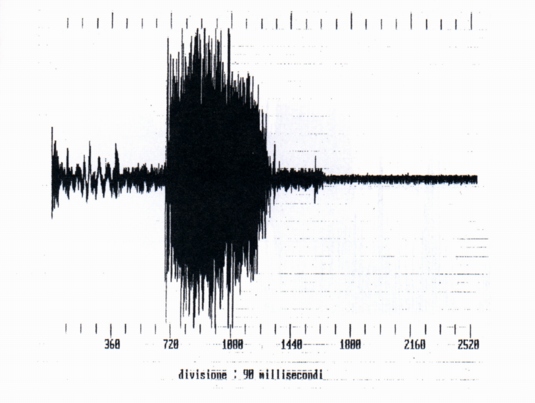
3.1- Single whistles (SW) (Fig. 1)
We have analysed ten whistles, of which four belonging with certainty to 3 different marmots, denominated 1, 2, 3; the other 6 whistles con be assigned either to the preceding marmots, and also to others. For only two whistles we have certain data about the nature of the phenomenon that precedes the emission, (ersatz of jay and of alpine chough). The length of the whistle is between 152 ms (ersatz of alpine chough) and 270 ms (jay' ersatz).
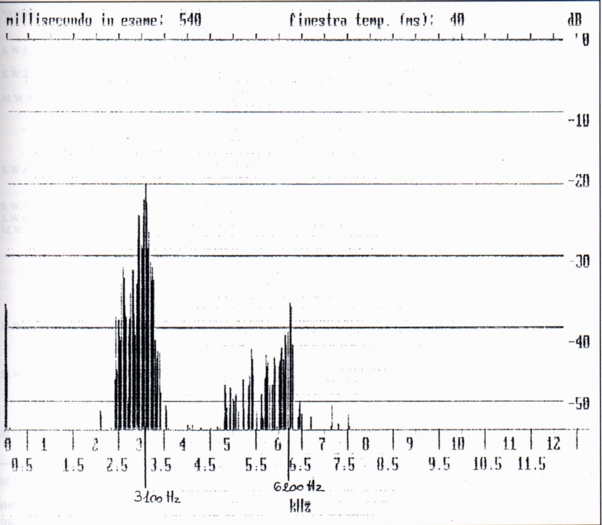
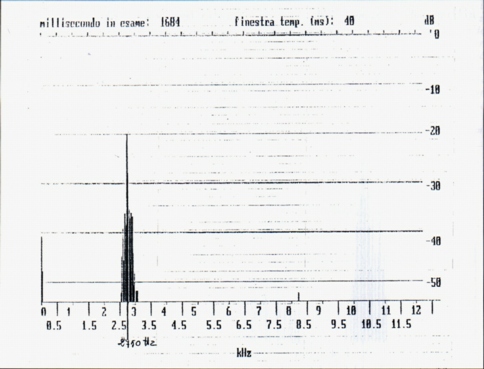
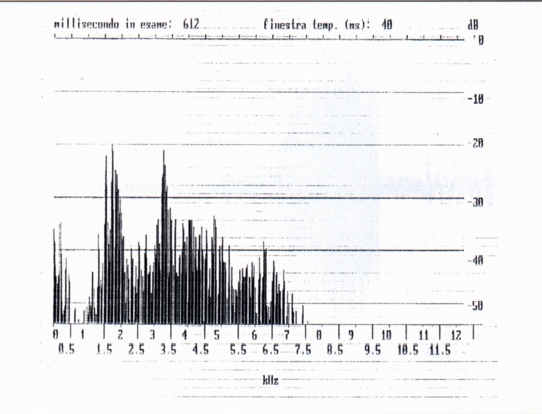
From the spectral point of view the single whistles are complex harmonic sounds of two or three components, the fundamental, the second and sometimes the third harmonic (Fig. 1a, Fig. 1b); in this they differ from human whistles (Fig. 2), where it is generally present the fundamental and the third harmonic; the variability of the spectrum shows a variability of shape of the front extremity of the vocal tract during the emission. The fundamental results between 2200 Hz (buzzard' and alpine chough' ersatz) and 3200 Hz (unknown stimulus). The frequency of the maximum of amplitude is always the fundamental, whistle the maximum frequency coincides with the frequency of the more elevated component (Tab. A).
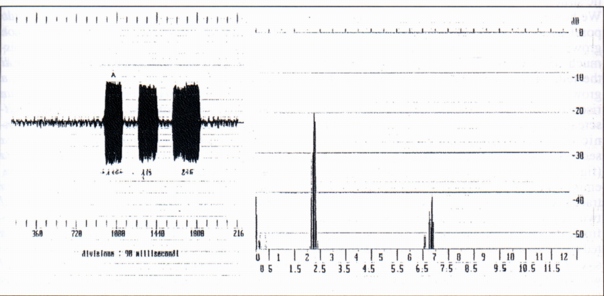
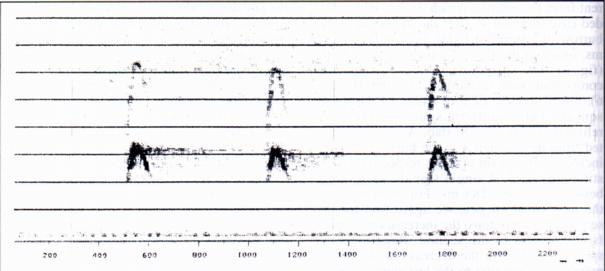
3.2- Multiple whistles (MW) (Fig. 3)
12 multiple whistles each constituted by a number of following whistles included between three and seven have been analysed; of the twelve multiple whistles, seven have certainly been emitted by 3 marmots called 1, 4, 6. For two multiple whistles and multiple alternate whistles to squeaks it is known the preceding physic phenomenon (buzzard' ersatz, a beam of light incident in the burrow); for the other 5 the whistle was emitted whistle in the area there was a fog-bank arising (even if we can't however assert that the fog was the cause of the emission). The duration of the following whistles are always different with irregular low, a part from the fact that the second whistle in nine cases resulted of lesser duration than the first. For the marmot 6 it was possible to record five multiple whistles (n° 13,14, 15, 16, 17), respectively constituted by 7, 3, 3, 3, 4 following whistles, therefore to establish, with "t" of Student, if the differences between the average duration of the single whistles were statistically significant. As a result the differences were for the most significant (from the comparison between whistles n°13 and 15, 15 and 16, 16 and 17). The differences between the whistles 13 and 14, 15 and 17 did not result significant. To these irregularities of the duration of the following whistles it is opposed the regular increase of the duration of the intervals of silence between the following single whistle (Fig. 3). Bearing in mind that the interval of silence between the following whistles corresponds to the inhalation [2], we can suppose that the regular increase of the intervals is bound to the process of nervous regulation of breathing (Hering and Brener's reflex) (Fig. 3). From the spectral standpoint, the spectra of the following single whistles constituent the multiple whistles contain from 2 to 4 harmonic components. The fundamental is included between 2200 and 3200 Hz (Tab. A). Considering again the case of the marmot 6 for which have been relieved 5 following series of multiple whistles, from the data relative to the loudest and to the weakest of each series, we observe that in 4 out of 5 cases, that is excluding the first series, the fundamental of the loudest is greater than that of the weakest, in accordance with known data relative to human vowels (Tab. A).
3.3- Squeak (single and multiple) (Fig. 4 and 5)
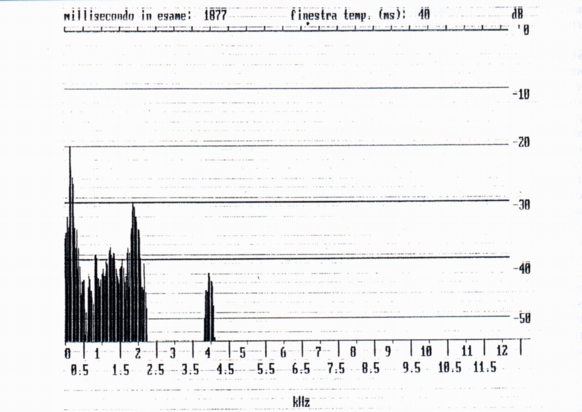
It is a signal with particular time wave different from the whistle, which, in the only recorded case (marmot 4, after the incidence in the burrow of a beam of light), is of duration 141 ms, lower to the field of values of the whistle (Fig. 4). The spectrum presents two spectral components with fundamental to 1900 Hz (Fig. 6), lower than the field of values of the single squeak. Three multiple squeaks have been relieved (n° 26, 27, 28) (Tab. A) (Fig. 5) emitted from the marmot 4 in the burrow after the incidence of a beam of light, respectively constituted by 2, 3, 4 single signals of duration between 102 and 283 ms. For what regards the interval of silence, we again find for the squeak of 3 emissions, the increase of duration from the first to the second. Regarding the spectra we observe the decrease of the fundamental from the first to the last signal (squeak 26, from 2550 to 1950 Hz; squeak 27, from 2700 to 1900 Hz; squeak 28, from 2200 to 1800 Hz). From such data it would result for the squeak a phonatory mechanism similar to that of the whistles, that is the vocal chord wouldn't be in vibration; some doubts however arise from examination of the spectra in the time of maximum amplitude, i.e. those reported in Fig. 4, between 0 and 200 Hz.
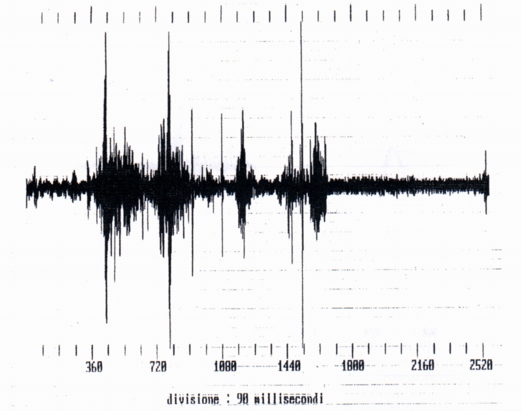
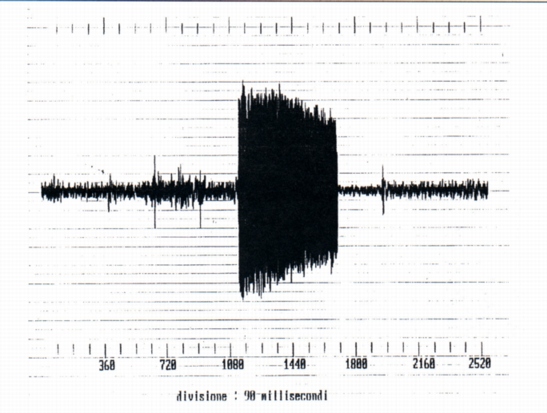
3.4- Growls and screams (Fig. 7 and Fig. 8)
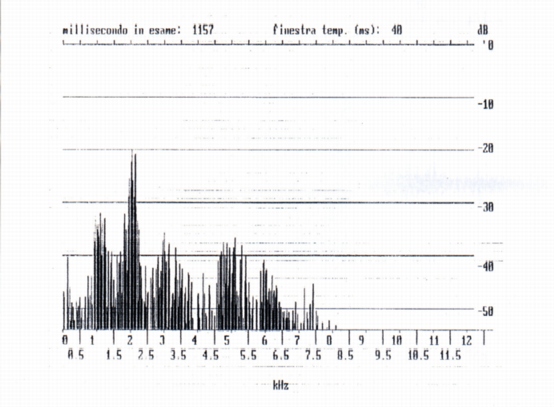
They are signals of temporal and spectral characters different from the whistles and squeaks, emitted during the capture of marmots 4 and 5. They are of duration clearly more elevated of the whistles and squeaks, that is 227 ms (growl, marmot 4) and 565 (scream, marmot 5) (tab. A). From a spectral point of view, that is timbric, they are "less clean" signal, a whole of sounds of harmonic structure to discreet components, with working vocal chords (similar to human vowels) and of noise, to continuous spectrum (due to the phenomena of viscous friction in the vocal tract, where the area of the section is lower). The consequence of that is that the following spectral maxima aren't exactly whole multiples of the fundamental (Fig. 9). The fundamental of the growl is included between 300 and 500 Hz (very lower that the fundamental of the single whistles) with other harmonic components and phormantic structure.
The fundamental of the scream (i.e. of the marmot 5) is value near to that of the growls with other harmonic component and phormantic structure.
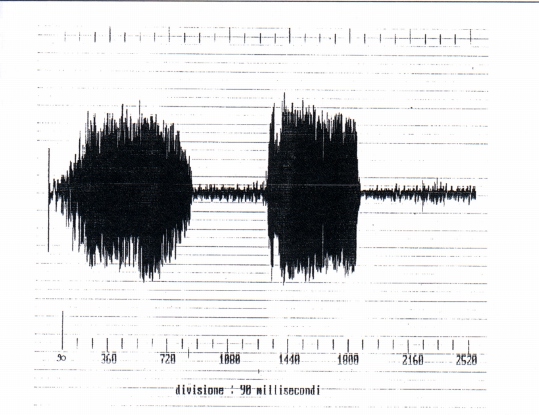
We however must observed that from the temporal point of view the screams differ to the growls because they have the staff and the end much more clear (Fig. 8). It must be relieved the temporal wave form is very irregular for growls and also for screams. From the comparison of the spectra of the growls and of the screams it results that for the first the harmonic of maximum amplitude is part of the second formant, for the screams of the third (Fig. 10). From which derives that during the emission of the scream the shape of the vocal tract is different from that of the growl. In the two cases examined the fundamentals are the first inferior to 500 Hz, but non-easily evaluated (for the presence of noise that is part of the signal), the second equals to 200 Hz about. It was also analyzed a mixed signal (Fig. 1l) constituted by a growl and a scream following of duration 758 and 566 ms respectively, with 448 ms of interval of silence. From the spectral point of view the two signals have numerous formants with fundamental frequencies nearly 300 and 200 Hz respectively.
4- SUMMARY
From the previous exposure results that, in spite of the limited number of emissions which follow with certitude determinate stimuli, at least as temporal succession, to stimuli constituted from ersatz of alpine chough and of jay follow single whistles, to buzzard ersatz multiple whistles; whereas to a beam of light incident in the burrow follow simple and multiple squeaks. The capture is on the contrary accompanied from screams and growls following.
We have observed that from the physic point of view the difference between whistle from one part, and screams and growls from the other, is in the fact that the firsts are signals at harmonic structure devoid of noise, whereas in the second the noise "pollutes" the signal.
To the contraposition of physic type could correspond the "semantic" contraposition between a whistle that convery information and screams and growls that, from a anthropomorphic point of view, express a emotional reaction to an event that directly regards the animal body.
retour/back
ANALISI PRELIMINARE DEI PARAMETRI ACUSTICI TEMPORALI E SPETTRALI DI EMISSIONI SONORE DI MARMOTA MARMOTA
Bassano°, Durio P.°, Facette C.*, Macchi E.°, Piazza R.*, Trinchero C.*
° Dipartimento di Produzioni Animali, Epidemiologia ed Ecologia - Università di Torino. Via Nizza 52, 10126 Torino, Italy
* Dipartimento di Morfofisiologia Veterinaria - Università di Torino. Via Campana 16, 10126 Torino, Italy
Â
1 - Introduzione
Dalla letteratura riguardante le emissioni sonore delle marmotte (Lenti Boero D., 1988-89), in particolare i segnali d'allarme, risulta che, dal punto di vista della motivazione, essi vanno studiati in base a vari modelli interpretativi, fra i quali si ricordano quelli che si basano sulle seguenti ipotesi:
a) l'ipotesi semantica, che implica il fatto che nell'animale esiste una forma di rappresentazione del monde esterno, dal quale arrivano uno o più segnali a cui segue temporalmente una risposta comportamentale;
b) l'ipotesi dell'eccitamento, per cui nel segnale emesso dall'animale viene codificato un particolare stato emotivo "determinato" dal contesto esterno;
c) l'ipotesi dell'inganno con due possibilità per l'animale: 1) segnalare un predatore inesistente allo scopo di approfittare della fuga generale per appropriarsi di risorse incustodite; 2) non segnalare un predatore esistente permettendo che esso agisca indisturbato. Nel lavoro (Noyes D.H e Holmes W.G., 1979) si descrive l'osservazione del diverso comportamento delle marmotte in relazione alla maggiore o minore gravità di rischio. Lo stimolo comportamentale era costituito da una sagoma d'aquila a comando radio.
à evidente che la risposta al quesito della motivazione, in varie situazioni contestuali, richiede uno studio dettagliato delle caratteristiche fisiche delle emissioni, ripetuto per ciascun tipo di emissione, in modo che l'analisi dei parametri in relazione ai dati del contesto possa dare una risposta in termini di analisi statistica multivariata, sulla esistenza o non esistenza di un rapporto causa/effetto fra il segnale esterno (o l'insieme dei segnali esterni) ed il comportamento del soggetto, definito dall'insieme dei valori di un certo numero di parametri acustici del segnale. La presente ricerca vuol fornire un contributo in tal senso, nel caso delle emissioni di Marmota marmota. In (Lenti Boero D., 1988-89) troviamo per Marmota marmota la descrizione dei seguenti tipi di richiami d'allarme: Fischio Multiplo (FM), definito come "suono acuto consistente in una serie di segnali elementari ad alta frequenza, emessi in successione più o meno rapida", rilevato quando si avvicinano animali terrestri o uomini; Fischio Singolo (FS), segnale unico che "non viene ripetuto", emesso in occasione del passaggio aereo di un potenziale predatore; Fischio Multiplo rallentato (FMR), definito come segnale "più simile ad uno squittio che ad un vero fischio" di tonalità più bassa, di motivazione non ancora chiarita; nella presente relazione sarà chiamato "squittio".
Guaito, suono "simile al guaito di un cane "; Ringhio, suono "simile al ringhio di un cane"; Tooth chatter, (Waring G., 1966) suono ottenuto facendo battere rapidamente gli incisivi; Grida (Waring G., 1966), emissioni ottenute prolungando fischi singoli.
Nella presente relazione si sono chiamate "grida " i particolari segnali sonori emessi durante la cattura. Nella presente ricerca sono stati analizzati fischi singoli e multipli, squittii, ringhi, grida, oltre all'insieme del segnali emessi durante e dopo la cattura.Dal punto di vista fonatorio, va ricordato che, mentre per i fischi singoli e multipli (e probabilmente per gli squittii) le corde vocali non sono in vibrazione, per gli altri segnali si ha vibrazione. A tale conclusione si arriva in base al dati rilevati della frequenza fondamentale del segnali sonori mediante confronto fra le dimensioni del condotti vocali dell'uomo e delle marmotte, dimensioni che possono essere considerate circa inversamente proporzionali alla frequenza fondamentale media.
2- Materiali e metodi
Le vocalizzazioni di Marmota marmota sono state registrate con l'impiego di un registratore Uher 4400 dotato di microfono unidirezionale e parabola.Le registrazioni sono state effettuate in due colonie poste a quote differenti (1700 e 1740 m s.l.m.) all'interno ed ai confini del Parco Naturale Orsiera Rocciavré (Alpi Occidentali) ed in una colonia presente nel Vallone dell'Ischiator (1791 m s.l.m.), Valle Stura (Provincia di Cuneo - Alpi Occidentali).
Nelle prime due zone di studio (Parco Orsiera), allo scopo di stimolare la reazione vocale delle marmotte, sono state utilizzate sagome di diverse specie di Uccelli, rapaci e non (Aquila reale -Aquila chrysaetos-, Poiana -Buteo buteo-, Corvo imperiale -Corvus corax-, Falconidae, Gracchio -Pyrrhocorax graculus, Ghiandaia -Garrulus glaudarius-), che venivano fatte cadere lungo un cavo sospeso 20 metri sopra le tane delle Marmotte.
Per rendere possibile tale operazione si sono scelte due colonie poste alla base di pareti rocciose, dalla cui sommità venivano rilasciate le sagome.La lunghezza del cavo sospeso era rispettivamente di 180 e 250 metri.Il punto di partenza delle sagome doveva essere celato alla vista delle Marmotte. Gli animali infatti che vedevano l'operatore rilasciare la sagoma non hanno mai emesso segnali acustici. Emissioni sonore sono state anche emesse durante l'improvvisa comparsa di nebbia.
Nella terza zona di studio (Valle Stura), si sono invece registrate vocalizzazioni di animali catturati con trappole e manipolati ed emissioni sonore provocate dal passaggio di cani e persone, che pero non sono risultate tecnicamente utilizzabili. Le analisi fisiche delle emissioni sono state effettuate mediante un Digital Sonagraph 7800 collegato mediante interfarcia 7812 - 488 ad un PC IBM compatibile dotale di programmi atti a fornire andamenti temporali con diverse scale, sonogrammi dei segnali con frequenza massima sia 6400 sia 12800 Hz e spettri sia con intervalli temporali regolabili da 5 a 160 ms, sia globali per intervalli prefissati fino a 2500 ms.
3 - Analisi sperimentali
(singoli e multipli), squittii (singoli e multipli), ringhi e grida. Poiché gli ultimi due segnali sono stati rilevati durante la cattura e si hanno emissioni di grida alternate a ringhi, i due segnali saranno trattati insieme.
3.1 - Fischi singoli (FS) (Fig. 1)
Sono stati analizzati dieci fischi, di cui quattro attribuibili con certezza a 3 diverse marmotte, denominate 1, 2, 3; gli altri 7 fischi possono essere attribuiti sia alle marmotte precedenti, sia ad altre. Per soli due fischi si hanno dati sicuri sulla natura del fenomeno che precede l'emissione, (sagoma di ghiandaia e di gracchio). La durata del fischio é compresa fra 152 ms (sagoma di gracchio) e 270 ms (sagoma di ghiandaia).
Dal lato spettrale, i fischi singoli sono suoni complessi armonici a due o tre componenti, la fondamentale la seconda e talvolta la terza armonica (Fig. 1a, Fig. 1b); in ciò differiscono dai fischi umani (Fig. 2), dove é generalmente presente la fondamentale e la terza armonica; la variabilità di spettro indica una variabilità di forma della estremità anteriore del condotto vocale durante la emissione. La fondamentale risulta compresa ira 2200 Hz (sagome di poiana e di gracchio) e 3200 Hz (stimolo ignoto).
La frequenza del massimo di ampiezza é sempre la fondamentale, mentre la frequenza massima coincide con la frequenza della componente più elevata. (Tab. A).
3.2 Fischi multipli (FM) (Fig. 3)
Sono stati analizzati 12 fischi multipli costituiti ciascuno da un numero di fischi successivi compreso fra tre e sette; dei dodici fischi multipli, 7 sono stati certamente emessi da 3 marmotte denominate 1, 4, 6.
Per due fischi multipli e fischi multipli alternati a squittii è noto il fenomeno fisico precedente (sagoma di Poiana, fascio luminoso incidente nella tana); per altri 5 il fischio è stato emesso mentre nella zona saliva un banco di nebbia (anche se non possiamo comunque affermare che la nebbia sia stata la causa dell'emissione).
Le durate dei fischi successivi sono sempre diverse con legge irregolare, a parte il fatto che il secondo fischio per nove casi è risultato di durata inferiore al primo.
Per la marmotta 6 è stato possibile registrare cinque fischi multipli (n°13, 14, 15, 16, 17), costituiti rispettivamente da 7, 3, 3, 3, 4 fischi successivi, quindi stabilire, col "t" di Student, se le differenze fra le durate medie dei fischi singoli erano statisticamente significative.
Come risultato le differenze sono risultate in maggior parte significative (confronti fra fischi n° 13 e 15, 15 e 16, 16 e 17).Non sono risultate significative le differenze fra i fischi 13 e 14, 15 e 17). A queste irregolarità delle durate dei fischi successivi si contrappone il regolare aumento della durata degli intervalli di silenzio fra i successivi fischi singoli (Fig. 3).
Tenendo presente che l'intervallo di silenzio tra i fischi successivi corrisponde all'inspirazione [2], si può ipotizzare che il regolare aumento degli intervalli sia legato ai meccanismi di regolazione nervosa del respiro (riflesso di Hering e Breuer) (fig. 3). Dal lato spettrale, gli spettri dei successivi fischi singoli costituenti il fischio multiplo contengono da 2 a 4 componenti armoniche. La fondamentale è compresa fra 2200 e 3200 Hz. (Tab. A). Considerando di nuovo il caso della marmotta 6 per la quale sono stati rilevate 5 successive serie di fischi multipli, dai dati relativi al più forte ed al più debole di ciascuna serie, si osserva che in 4 casi su 5, ossia escludendo la prima serie, la fondamentale del più forte è maggiore di quella del più debole, in accordo con noti dati relativi alle vocali umane. (Tab. A)
3.3 - Squittio (singolo e multiplo) (Fig. 4 e Fig. 5)
à un segnale con andamento temporale particolare diverso dal fischio, il quale nell'unico caso registrato (marmotta 4, dopo l'incidenza sulla tana di un fascio luminoso), è di durata 141 ms, inferiore al campo di valori del fischio (Fig. 4).Lo spettro presenta due componenti spettrali con fondamentale a 1900 Hz (Fig. 6), più bassa del campo di valori dello squittio singolo.
Squittii multipli (n° 26, 27, 28; tab. A) (Fig. 5) emessi dalla marmotta 4 nella tana dopo incidenza di fascio luminoso, costituiti rispettivamente da 2, 3, 4 segnali singoli di durata fra 102 e 283 ms. Per ciò che riguarda l'intervallo di silenzio, si ritrova per la squittio a 3 emissioni, l'aumento di durata dal primo al secondo. Per ciò che riguarda gli spettri si osserva la diminuzione della fondamentale dal primo all'ultimo segnale (squittio 26, da 2550 a 1950 Hz; squittio 27, da 2700 a 1900 Hz; squittio 28, da 2200 a 1800).
Da tali dati risulterebbe per la squittio un meccanismo fonatorio analogo a quello dei fischi, ossiale corde vocali non sarebbero in vibrazione; alcuni dubbi peraltro derivano dall'esame degli spettri nell'istante di massima ampiezza, ad esempio quelli riportati in Fig. 4, fra 0 e 200 Hz.
3.4- Ringhi e grida (Fig. 7 e Fig. 8)
Sono segnali di caratteri temporali e spettrali diversi dai fischi e dagli squittii, emessi durante la cattura delle marmotte 4 e 5.
Essi sono di durata nettamente più elevata dei fischi e degli squittii, ossia 227 ms (ringhio, marmotta 4) e 566 (grido, marmotta 5) (tab. A).
Dal punto di vista spettrale, ossia timbrico, sono segnali meno "puliti", insieme di suoni di struttura armonica a componenti discrete, con corde vocali in funzione (analoghi alle vocali umane) e di rumore, a spettro continuo (dovuto a fenomeni di attrito viscoso nel condotto vocale, dove l'area della sezione è minore). La conseguenza di ciò è che i successivi massimi spettrali non sono esattamente multipli interi della fondamentale (Fig. 9). La fondamentale del ringhio è compresa tra 300 e 500 Hz (mollo più bassa della fondamentale dei fischi singoli) con altre componenti armoniche e struttura formantica.
La fondamentale del grido (ad esempio della marmotta 5) è di valore vicino a quella dei ringhi con altre componenti armoniche e struttura formantica. Va inoltre osservato che le grida dal punto di vista temporale differiscono dai ringhi avendo l'attacco e la fine assai più netto (Fig. 8).
Va rilevato che la forma d'onda temporale è assai irregolare sia per 1 ringhi sia per le grida.
Dal confronto degli spettri dei ringhi e delle grida risulta che per i primi l'armonica di massima ampiezza fa parte della seconda formante, per le grida della terza, (Fig. 10). Da ciò deriva che durante l'emissione del grido la forma del condotto vocale è diversa da quella del ringhio.
Nei due casi esaminati le fondamentali sono la prima inferiore a 500 Hz, ma difficilmente valutabile (per la presenza di rumore facente parte del segnale), la seconda pari circa a 200 Hz.
à stato inoltre analizzato un segnale misto (Fig. 11) costituito da un ringhio e un grido successivi di durata 758 e 566 ms rispettivamente, con 448 ms di intervallo di silenzio. Dal punto di vista spettrale i due segnali hanno numerose formanti con frequenze fondamentali circa 300 e 200 Hz rispettivamente.
4 - Conclusioni
Dall'esposizione precedente risulta che, malgrado il numero limitato di emissioni che seguono con certezza determinati stimoli, almeno come successione temporale, a stimoli costitutivi da sagome di Gracchio e di Ghiandaia seguono fischi singoli, a sagoma di Poiana fischi multipli; mentre ad un fascio di luce incidente nella tana seguono squittii, semplici e multipli. La cattura è invece accompagnata da grida e ringhi successivi. Si osserva che dal punto di vista fisico la differenza tra fïschi da una parte, e grida e ringhi dall'altra, sta nel fatto che i primi sono segnali a struttura armonica privi di rumore, mentre nei secondi il rumore "inquina" il segnale. Alla contrapposizione di tipo fisico potrebbe corrispondere la contrapposizione "semantica " tra fischio che trasmette informazioni e grida e ringhi che, dal punto di vista antropomorfico, esprimono una risposta emotiva ad un evento che riguarda direttamente il corpo dell'animale.
retour/back
Â
Â
Â
Â
Â
Â
Â
Table A: VOCAL. = VOCALISATION - STIM. = STIMULUS - S.W. = SINGLE WHISTLE - M.W. = MUTIPLE WHISTLES - S.S. = SINGLE SQUEAK - M.S. = MULTIPLE SQUEAKS - G. = GROWL - SCR. = SCREAM - CAPT. = CAPTURE - Fo = FUNDAMENTAL FREQUENCY
NOYES D.H., HOLMES W.G., 1979 - Behavioural responses of free-living Hoary marmots to a model golden Eagle. Joumal of mammology, vol. 60, n°2: 408-411.