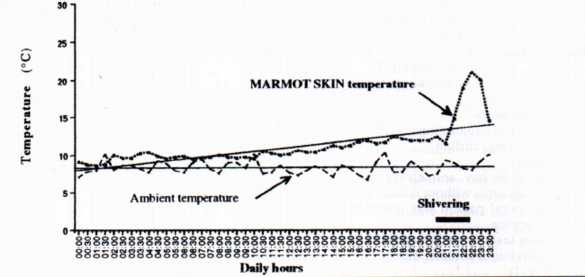
Fig. 2: the evolution of marmot skin temperature during one of rewarming phases. Non shivering thermogenesis and shivering seemend to be used for the increase of the body temperature.
Cochet N., Le Berre M., Le Guelte L.
Laboratoire de Socioécologie et d'Ecoéthologie, Université Claude Bernard Lyon 1, F69007 Lyon, France.
Abstract - Three adult Alpine Marmots (Marmora marmota) were studied, during winter 91, in a dark room under constant ambient temperature (8°C). Behaviour and skin temperature were continuously recorded, during the whole period. Behavioural patterns and percentage of activity, observed during arousal phases, showed no significant variations from one arousal to another whatever the duration of the arousals may be. Constant ambient conditions allowed us to show a free-running circadian activity rhythm, the period of which varied from 25 to 34 hours, depending on the individuals. Marmots seemed able to adjust their activity in relation to duration of their arousals. Adjustment of activity and adoption of postures reducing heat losses might contribute to a minimization of energetic expenses.
INTRODUCTION
Hibernation is at present being studied, by many French researchers, but Alpine Marmot model seems to be discarded, despite its important sample in France.
Dubois published the first study on physiology of hibernating alpine marmot, at the end of the last century (1896).
Later, in the sixties and seventies, Kayser (1963) and Malan et al. (1973), studied some respirometric and energetic aspects on hibernating marmots. Very few works about behaviour and chronobiology have been realized, during hibernation, despite the fact that physiological aspects of hibernation of Alpine Marmot have often been studied.
Arnold (1988) showed energetic consequences of behavioural and social thermoregulation in family groups of hibernating Alpine Marmots. These two topics were (during winter 1991) the subject of the present study.
The aim was first, to describe behavioural patterns and temporal distribution of activity during arousal states and, second, to show up the possible existence of a free-running circadian activity, in relation to the constant darkness in which Marmota marmota lives in its natural habitat, during hibernation season.
METHODS
Three adult Alpine Marmots were used in this study, two males (called V and C) and one female (called L). They came from the Natural Park of Mercantour, in the department of Alpes Maritimes, France, and lived in captivity at the National Veterinary School of Lyon, since three years. These marmots belonged to different social groups. Two of them lived together in the same cage. Each marmot could be identified by its particular fur pigmentation. These three individuals were grouped in the same glazed cage, without food and water and placed in a cold dark room, under constant ambient temperature of 8°C.
Animal behaviour was continuously filmed and recorded by means of infrared camera and a time-lapse videotape recorder. Date and hour were recorded too. Temperature picks up, located in the false floor of the cage, allowed us to follow evolution of skin temperature of marmots during the season and precise arousal states.
Videotapes were analysed in order to draw up an inventory of different behavioural patterns and to establish the temporal distribution of activity during arousal phases. Frequencies and durations of behavioural patterns were noted for each individual. During each arousal, every behavioural pattern called "Sequence Motrice Simple" (S.M.S.) (Gervet & Truc, 1974) was expressed in percent of total number of S.M.S.. Daily durations of activity, expressed in minutes per hour, allowed us to realized actograms and periodograms, in order to define the periodicity of a possible activity rhythm.
STATISTICS
Behavioural patterns were compared on different non independant periods (periodical arousals). As we disposed of a limited number of animals Friedman non parametric statistical test was used with a threshold of 0.05. For statistical treatments of circadian rhythms in order to show a possible significant period of activity, Chi-square periodograms of Sokolove & Bushell (1978) were used.
RESULTS
Behavioural study
Marmots were observed during 91 days. The behavioural patterns were counted, mainly during arousal states. They were classified according to their affinity.
Social behaviours
- Allogrooming: one individual grooms the fur of another individual.
- Game: only observed between the two males at the end of the season, animals pushing themselves with their forelegs.
Cleaning behaviours
- Grooming: animals groom their fur.
- Nest building: animals reorganize the hay around themselves.
Others behaviours
- Displacements, explorations
- Jumps/shivering, curling
- defecations
- Chewing of the hay: animals split up the hay into little fragments without consumption of it.
This behavioural pattern was observed at the end of the season.
A Friedman test shows no significant evolution of behavioural patterns during hibernation season (Fig. 1)
Evolution of skin temperature
During the season, one of the three marmots awaked completely on one of the six temperature picks-up. Its skin temperature evolved according to two stages (Fig. 2). The first one showed a slow increase of skin temperature (rewarming speed was around 0.4°C/hours), probably initiated by utilization of brown fat. The second stage showed an acceleration of the rewarming process (speed was around 5°C/hours) which was related to shivering of the animal.
Arousals and activity
Mean duration of arousals was around 50 hours (Fig. 3) and was significantly different from one arousal to another (Friedman, p<0.05). Mean duration of hibernation phasis was about l150 hours (Fig. 3) and was significantly different from one arousal to another (Friedman, p<0,05). Mean duration of activity during arousal states, was expressed in percent of total awake time. Marmots were active only 12 to 16% of their awake time (Fig. 4). An interesting fact is that the percentage was not significantly different from one arousal to another whatever the duration of the arousal may be (Friedman, p> 0.7).
Rhythmicity study
During this experimentation, two rhythmicities were shown up:
- Free-running circadian rhythmicity related to the constant environmental factors of the room. The durations of marmot activity, obtained through analysis of videotape recording, allowed us to realize actograms for each animal (Fig. 5a).
In a first time, "full length" actograms were first realized, including the whole season of hibernation (91 days) with arousal and hibernation phases. Analyses of these data by Chi-square periodograms showed no evidence of significant period of activity rhythm. The hibernation phases, during which no activity was recorded during several consecutive days, created a "dilution effect" in the data. For that reason, they were deleted and data on arousal phases were joined end to end. "Rectified" actograms resulting from their transformations, allowed us to emphasize a temporal organization of activity (Fig. 5b).
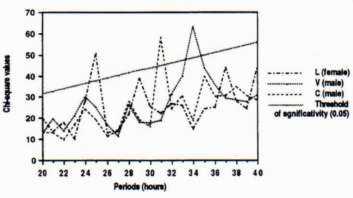
The rate of activity was more important at the beginning and at the end of each day. Chi-square periodogram analyses showed existence of two parts in these new actograms. A first part in which no significant period of activity rhythm can be shown. A second part, in which a significant period of activity was shown for each of the three individuals (Fig. 6). An important interindividual variability was observed between the three marmots. Periodicity was about 25 hours for the female, 31 hours for the C male and 34 hours for the V male.
DISCUSSION
Behavioural study shows that our marmots presented the same percentage of activity during awake phases. They seem to be able to adjust their rate of activity according to the duration of arousals. Studies of time-distribution confirm the functioning of the biological clock during hypothermic phases and the conservation of a circadian rhythm from one arousal to another. This fact was already demonstrated by Canguilhem (1991) in other species of hibernating rodents. The present study also shows a great variability of the free-running circadian rhythm between individuals. The difference between the significant period of L and V (9 hours) seems very important. No obvious reason can be given to explain such variability, observed on individuals submitted to the same experimental conditions. Variability might result from the influence of all the factors, which contribute to diversify populations. Variability could result too from data modifications required for the statistical treatment.
These hypotheses could be applied too, to the durations of arousals and of hibernation phases. In spite of similar experimental conditions, the data given by Arnold (1988) differ greatly from ours: torpor duration of his animals was around 344 hours (twice the values we observed) and awake duration was around 24 hours (half the values we observed).
CONCLUSIONS
Marmota marmota seems able to control its energetic potential by its behaviour. Self-management of its rate of activity and adoption of postures reducing heat losses (curling) might contribute to an important limitation of energy consumption during the arousal states.
ACKNOWLEDGEMENT
This work is part of the research program "Dynamics of space occupation by Alpine Marmot" supported by the French Ministery of Environment (EGPN 90-294). We would like to express our gratitude to Mme Chantal Jacquet of National Veterinary School of Lyon (INSERM U 271) for the loan of her three Alpine Marmot actresses and to Dr R. Allemand (UA CNRS 243) of Claude Bernard University of Lyon for his contribution to the statistical treatments of circadian activity rhythms.
RITMI CIRCADIANI E COMPORTAMENTO DI UN GRUPPO DI MARMOTTE ALPINE (Marmota marmota) IN IBERNAZIONE.
Cochet N., Le Berre M., Le Guelte L.
Laboratoire de Socioécologie et d'Ecoéthologie, Université Claude Bernard Lyon 1, F69007 Lyon, France.
Introduzione
Attualmente l'ibernazione viene studiata da numerosi ricercatori francesi, ma la Marmotta alpina, come modello di studio, sembra essere stato dimenticato, nonostante la sua notevole importanza in Francia.
Dubois pubblico il primo studio sulla fisiologia delle marmotte alpine in ibernazione alla fine del secolo scorso (1896). Più tardi, negli anni '60 e '70, Kayser (1963) e Malan (Malan et al., 1973), studiarono alcuni aspetti respirometrici ed energetici delle Marmotte in letargo. Sono stati realizzati pochissimi lavori sul comportamento e sulla cronobiologia durante l'ibernazione, nonostante che gli aspetti fisiologici dell'ibernazione siano stati spesso indagati. Arnold (1988) ha messo in evidenza le conseguenze energetiche della termoregolazione comportamentale e sociale di marmotte ibernanti in gruppi familiari. Questi due aspetti sono stati studiati durante l'inverno 1991. Lo scopo principale era quello di descrivere i moduli comportamentali e la distribuzione temporale delle attività durante gli stati di veglia e nel contempo di mettere in evidenza la presenza di una attività circadiana libera, correlata al buio costante in cui Marmota marmota si viene a trovare all'interno delle tane naturali durante la stagione di ibernazione.
Metodi
Sono stati utilizzati per questo studio tre adulti di Marmotta alpina, due maschi (chiamati V e C) e una femmina (chiamata L). Provenivano dal Parco Nazionale del Mercantour, nel Dipartiniento delle Alpi Marittinie, e sono vissute negli ultimi tre anni in cattività presso la Scuola Veterinaria Nazionale di Lione. Queste marmotte appartenevano a differenti gruppi sociali. Due di esse erano vissute nella stessa gabbia.
Ciascuna marmotta era riconoscibile dal colore della pelliccia. Questi tre individui sono stati posti in una gabbia di vetro, senza cibo né acqua in una stanza buia e fredda, a temperatura costante di 8°C. Il comportamento degli animali era costantemente filmato e registrato da una videocassetta ad infrarossi, con un videoregistratore a tempo, su cui venivano registrati anche data e ora del giorno. Sensori per la temperatura, sistemati in un sottofondo della gabbia, permettevano di seguire l'evoluzione della temperatura cutanea delle marmotte, durante il letargo e definire gli stati di veglia. Le registrazioni video sono state analizzate al fine di tracciare un inventario dei differenti moduli comportamentali e di stabilire la distribuzione temporale dell'attività durante le fasi di veglia. La frequenza e la durata dei moduli comportamentali sono stati registrati per ciascun individuo. Durante ogni veglia, ogni modulo comportamentale, chiamato "frequenza Motrice Semplice" (S.M.S. ) (Gervet e Truc, 1974) é stata espressa come percentuale sul numero totale di S.M.S. La durata giornaliera delle attività , espresse in minuti all'ora, ha permesso di realizzare degli actogrammi e dei periodogrammi al fine di definire la periodicità di un possibile ritmo di attività .
Metodi statistici
I moduli comportamentali sono stati confrontati tra periodi non indipendenti (risvegli periodici). Poiché si aveva a disposizione un numero limitato di animali si é usato il test statistico di Friedman, con una soglia di 0.05. Per il trattamento statistico dei ritmi circadiani, al fine di mettere in evidenza possibili periodi significativi di attività , é stato utilizzato il periodogramma Chi-quadro di Sokolove e Bushell (1978).
Risultati
Studio comportamentale
Le marmotte sono state osservate per 91 giorni. Sono stati definiti dieci moduli comportamentali, in particolare durante gli stati di veglia. Sono stati classificati in base alla loro affinità . Comportamenti sociali
- Allogrooming: un individuo pulisce la pelliccia di un'altro individuo.
- Gioco: osservato unicamente tra i due maschi verso la fine della stagione, gli animali si spingevano con le zanipe anteriori.
Comportamenti di pulizia
-Grooming: gli animali si puliscono la pelliccia.
-Costruzione del nido: gli animali risistemano la paglia attorno a loro.
Altri comportamenti
-Spostamenti, esplorazioni
-Salti/brividi/raggomitolamenti
-Defecazione
-Masticazione della paglia: gli animali trinciano la paglia senza cibarsene. Questo modulo comportamentale é stato osservato alla fine della stagione. Il test di Friedman mette in evidenza la mancanza di una significativa evoluzione dei moduli comportamentali durante la stagione di ibernazione (Fig. 1).
Evoluzione della temperatura cutanea
Nella fase di letargo, uno dei tre animali si sveglio completamente, nel corso di uno dei soi picchi di temperatura. La sua temperatura cutanea ebbe un andamento a due stadi (Fig. 2). Nel primo si é osservato un lento incremento della temperatura cutanea (velocità di riscaldamento: circa 0.4°C/ora), innescato probabilmente dall'utilizzazione del grasso bruno. Il secondo stadio presentava un accelerazione del processo di riscaldamento (la velocità era di circa 5°C/ora) in concomitanza di brividi.
Risveglio e attivitÃ
La durata media del risveglio era di circa 50 ore (Fig. 3), significativamente differente tra un risveglio e l'altro (Friedman, p<0.05). La durata media delle fasi di ibernazione era di circa 150 ore (Fig. 3) ed era significativamente differente tra un risveglio e l'altro (Friedman, p<0.05). La durata media delle attività durante gli stati di veglia é stata espressa in percentuale sul tempo totale di veglia. Le marmotte erano arrive solo per il 12-16% del tempo (Fig. 4). Un fatto interessante é che tale percentuale non era significativamente differente tra un periodo di veglia e l'altro, qualunque ne fosse la durata (Friedman, p>0.7).
Studio della ritmicitÃ
Durante questa sperimentazione si sono evidenziate due ritmicità :
- Periodicità ritmica dei risvegli che avvenivano ogni 5 giorni
- Ritmicità circadiana libera collegata ai fattori ambientali costanti della stanza.
La durata delle diverse fasi di attività delle marmotte, calcolata analizzando le registrazioni video, ci ha permesso di tracciare gli actogrammi di ogni animale (Fig. 5a).
In un primo tempo sono stati realizzati degli actogrammi "full length" (91 giorni) comprendenti le fasi di sonno e di veglia. L'analisi di questi dati, per mezzo del periodogramma Chi-quadro, ha messo in evidenza la mancanza di significativi ritmi periodici di attività . Le fasi di ibernazione, durante le quali non venne registrata alcuna attività per parecchi giorni consecutivi, creano un "effetto diluizione " dei dati. Per ciô queste fasi vennero omesse e le veglie unite le une alle altre. Gli actogrammi "rettificati" cosi ottenuti hanno permesso di evidenziare un'organizzazione temporale delle attività (Fig. 5b).
Il tasso di attività era più alto all'inizio del giorno rispetto alla fine. L'analisi del periodogramma Chi-quadro ha messo in luce l'esistenza di due fasi all'interno di questi nuovi actogrammi. Nella prima non si evidenzia alcun periodo di attività ritmica. Nella seconda esiste invece un periodo di attività significativo, caratteristico per ciascuno dei tre individui (Fig. 6).
Nelle tre marmotte esiste una notevole variabilità soggettiva. La periodicità era infatti di circa 25 ore per la femmina, 31 per il maschio C e 34 ore per il maschio V.
Discussione
Lo studio comportamentale evidenzia che le nostre marmotte presentano la stessa percentuale di attività durante gli stati di veglia. Esse sembrano in grado di adattare il loro grado di attività a seconda della durata dei risvegli. Gli studi sulla distribuzione temporale confermano il funzionaniento dell'orologio biologico durante le fasi ipodermiche e la conservazione di un ritmo circadiano, tra un periodo e l'altro. Questo fatto era già stato dimostrato da Canguilhem (1991), in altre specie di roditori ibernanti. Questo studio mette inoltre in evidenza la grande variabilità dei ritmi circadiani liberi tra i diversi individui. La differenza tra i valori di periodicità di L e V (9 ore) sembra essere di grande importanza. Non si riesce a trovare una motivazione per tale variabilità , dal momento che gli individui erano sottoposti alle stesse condizioni sperimentali. La variabilità puo essere il risultato d'azione dei diversi fattori che concorrono a diversificare le popolazioni. Oppure puo essere il risultato delle variazioni dovute al trattamento statistico dei dati. Queste ipotesi possono altresi essere applicate alla durata delle fasi di veglia e di sonno. Nonostante le condizioni sperimentali simili, i dati forniti da Arnold (1988) differiscono notevolmente dai nostri: la durata del torpore nei suoi animali era di circa 344 ore (il doppio circa dei valori da noi osservati) e la durata dei periodi di veglia era di circa 24 ore (la metà dei valori da noi osservati).
Conclusioni
Marmota marmota sembra in grade di controllare il proprio potenziale energetico per mezzo di variazioni comportamentali. L'auto-controllo del proprio livello di attività e l'adozione di posture atte alla riduzione delle perdite di calore (acciambellarsi) possono contribuire in maniera importante alla limitazione del consumo energetico durante gli stati di veglia.
Ringraziamenti
Questo lavoro é parte del programma di ricerca "Dinamica dell'occupazione dello spazio della Marmotta alpina " finanziato dal Ministero dell'ambiente francese (EGPN 90-294). Voglianio esprimere la nostra gratitudine a Mme Chantal Jacquet, della Scuola Nazionale di Veterinaria di Lione, (INSERM U 271) per averci concesso le sue tre marmotte e al Dr.R. Allemand (UA CNRS 243), dell'Università Claude Bernard di Lione, per il suo contributo al trattamento statistico dei dati relativi ai ritmi circadiani di attività .
ritorno/back
REFERENCES
ARNOLD W. (1988). Social thermoregulation during hibernation in Alpine Marmots (Marmota marmota). J. Comp. Physiol B, 158: l5l-156.
CANGUILHEM B. (1991): Temporal organisation in hibemating rodens. 3rd Int. Coll. Rodens spatium, Lyon, March 1991 (in press).
DUBOIS R. (1896): Etudes sur le mécanisme de la thermogénèsc et du sommeil chez les mammifères: physiologie comparée de la marmotte. Ann. Univ. Lyon, 25.
GERVET & TRUC (1974): Les problèmes de l'analyse des enchaînements séquentiels d'actes moteurs. Bull. Int S.F.E.C.A. Rennes: p 9- Ill.
KAYSER C. (1963): Relation entre la température ambiante, la température centrale, le poids et la thermogénèse des hibernants en hibernation profonde. C.R. Sc. Mem. Soc. Biol., 157: 2065-2067.
MALAN A., ARENS H. ET WAECHTER A. (1973): Pulmonary respiration and acid-base state in hibernating marots and hamster. Respiration physiology, 17 : 45-61.
SIEGEL S. (1956): The Friedman two-way analysis of variance by ranks. In: Nonparametric Statistics for the behavioural sciences, Mac Graw Hill Edition, Tokyo, 166-173.
SOKOLOVE P.G. & BUSHELL B. (1978). The Chi-square periodogram: its utility for analysis of circadian rhtyhm. J. Theoret. Biol., 72: 131-1611.
ritorno/back
Fig. 1: Tests on the behavioural patterns realized by the three animals, on seven arousals (Friedman test, df = 6)
|
BehaviouralPatterns (SMS) |
Chi-Square value |
probability |
significativity |
|
Allogrooming |
12.35 |
0.1> p >0.05 |
N.S. |
|
Nest building |
3.53 |
p> 0.7 |
N.S. |
|
Displacements |
3.17 |
p> 0.7 |
N.S. |
|
Grooming |
9.39 |
p> 0.l |
N.S. |
|
Jumps |
7.46 |
p> O.2 |
N.S. |
|
Explorations |
10.07 |
p> 0.1 |
N.S. |
|
Curling |
6.5 |
p> 0.3 |
N.S. |
|
Defecations |
10.50 |
p> 0.l |
N.S. |
Fig. 2: the evolution of marmot skin temperature during one of rewarming phases. Non shivering thermogenesis and shivering seemend to be used for the increase of the body temperature.
Fig. 3: Mean durations of arousals and hibernation phases for the 3 animals. 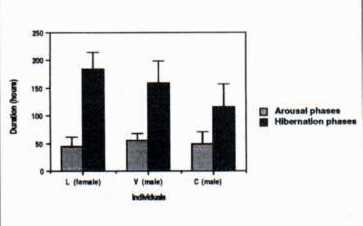 ritorno/back | Fig. 4: Mean activity (in % of total awake time) during arousals. 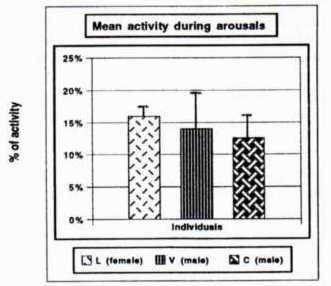 ritorno/back |
Fig. 5a: "Complete" actogram of the female
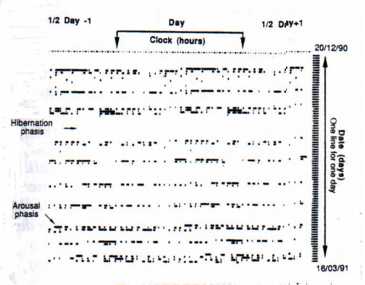
ritorno/back
Fig. 5b: "Rectified" actogram of the female

ritorno/back
Fig. 6: Periodograms of the 3 marmots realised from the second parts of the rectified actograms.
ritorno/back