MARMOTA CAMTSCHATICA PALL. - NEARCTIC ELEMENT IN PALEARCTIC MARMOTA FAUNA
Lyapunova E.A., Boyeskorov G.G., Vorontsov N.N.
N.K. Koltsov Institute of Developmental Biology, the USSR Academy of Sciences, Moscow; Institute of Biology, Sibirian branch of the USSR Academy of Sciences, Yakutsk; USSR Ministry of Natural Resourccs, Management and Environmental Protection, Moscow
Abstract - The authors studied the caryology of Holoartic marmots (M. b. bobac, M. caudata aurea, M. menzbieri zahidovi, M. baibacina centralis, M. s. sibirica, M. camtschatica bungei, M. camtschatica doppelmayeri and M. c. camtschatica). Chromosomes preparation were made using a standard technique of Ford and Hamerton. Palearctic species are homogenous while Nearctic ones show broad caryological divergence.
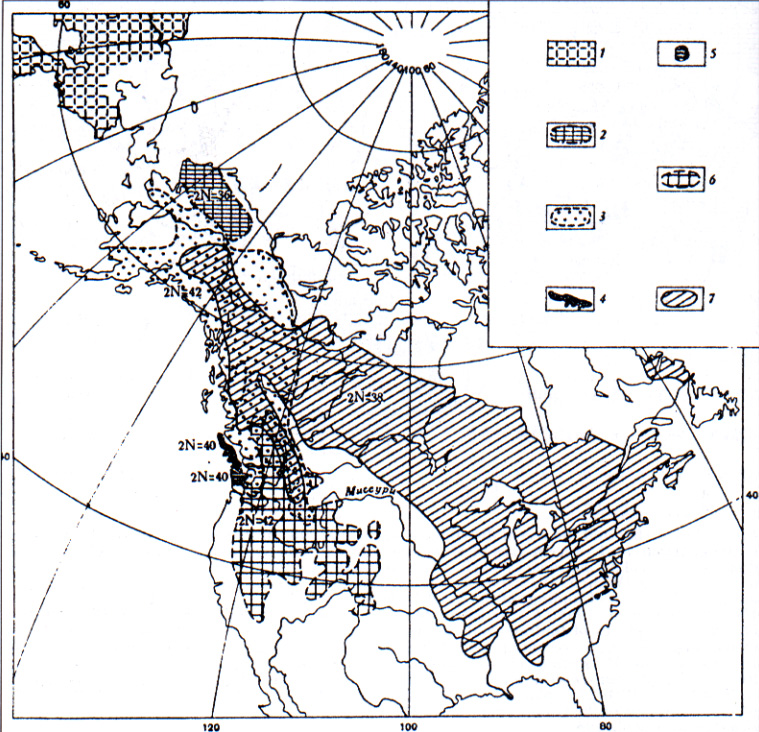
Comparative caryology of Holarctic marmots was studied in the sixties by routine chromosomal staining technique before discovering the methods of differential staining of chromosomes. In contrast with caryologically homogenous the most part of Palaearctic species (2n=38, NFa=64-66; Vorontsov et al., 1969; Lyapunova et al., 1969; Fernandez-Donoso et al., 1969), excluding M. camtschatica (2n=40, NFa=62; Lyapunova et al., 1969) (Fig. 1) nearctic species of Marmota have a broad caryological divergence: M. flaviventris and M. caligata - 2n=42, NFa=62; M. broweri -2n=36, NFa=62; M. monax has 2n=38, NFa; M. vancouverensis and M. olympus have 2n=40 (Rausch et al., 1965; Hoffmann et al., 1968). Sharp differences in the Ievels of the caryological divergence between the Palaearctic and Nearctic marmots correlated with strictly allopatry of the Palaearctic species (Fig. 2) and semisympatry of the Nearctic marmots (Fig. 3).
Sympatric character of distribution and divergence in chromosome numbers of the Nearctic species reflects more ancient hystory of the American forms as compared with the allopatric distribution and similarity of chromosomes of the Palearctic species (exduding M. camtschatica) (Vorontsov et al., 1969; Lyapunova et al., 1969, Vorontsov et al., 1976; Nadler et al., 1976).
Recent distribution of the most Palaearctic species has a relict character as a result of Holocaenic reducing of areas of the mountain species (M. marmota), steppen species (M. bobak), extinction (Marmota caucasica), restoration of the forests in postglacial time (West-Siberian populations of M. sibirica and M. camtschatica) and anthropogenic pressure. Distribution of M. camtschatica is quite similar with spreading of Spermophillus parryi in Siberia. M. camtschatica occupied mountain alpine area, and S. parryi distributed also in plain steppen-islands and steppen-tundra of the North-East of Siberia. Yet both species have a very mosaic pattern of the distribution, such mosaic distribution is more typical for M. camtschatica. In this species one population separates from another by many hundred kilometers. Morphological differences between M. c. camtschatica from Kamtchatka peninsula, M. c. bungei from North Yakutia and M. c. doppelmayeri from Transbaikalia are significant (Gromov et al., 1965) as well as ecological differences (Bibikov, 1965). Early we studied caryology of these marmots only from Kamtchatka population. It is necessary to support conspecificity of the different isolated subspecies of M. camtschatica.
This publication is a part of our project of the reinvestigation of cytogenetics of the Holarctic marmots by modern chromosomal technique.
MATERIALS AND METHODS
6 species of the palaearctic marmots were investigated karyologically.
Marmota bobak bobak Muller. The Eastern Ukraine, Lugansk district, "Streletskaya step" preserve 1 male;
M. caudata aurea Blanf. 15 specimens from the three point of the area:
1) The Pamir-Alay mountains, Gissar ridge, near Anzob pass - 4 males and 2 females;
2) The Eastern Pamirs. Murgab region, near Chichekty kishlak - 2 males, 4 females;
3) Central Thian Shan, Talass ridge, near Besh-Tash pass - 1 male, 2 females.
M. menzbieri zahidovi Petrov. Western Thian Shan, Karamin ridge, the border of Angren plateau - 1 male, 1 female.
M. baibacina centralis Thomas. Eastern Thian Shan, Trans-Ili Ala-Tau - 1 male, 1 female.
M. sibirica sibirica Radde. Transbaikalia, Chita distr., vicinity of the Borzya railway station - 1 male.
M. camtschatica bungei Kastsch. North Yakutia: 1. Kharaulakh mountains, eastern bank of the Lena river - 1 male; 2. Verkhoyansky ridge, the upper reaches of the Kele river - 2 males.
M. camtschatica doppelmayeri Birula. Buryatia, Barguzin ridge, the upper reaches of the Urbikan river - 1 female.
M. camtschatica camtschatica Pall. Kamtchatka, the slope of Avacha volcano - 1 female.
Chromosomes preparation were made using a standard technique of Ford and Hamerton (1956). Chromosomes of two males M. camtschatica bungei from Verkhoyansky ridge were studied with AgNOR-staining in modification of Howell & Black (1980).
RESULTS AND DISCUSSION
As our works on Marmota (Vorontsov et al., 1969; Lyapunova et al., 1969) were published only in Russian, we include in this report all the materials we have at our disposal. The new data we received on M. c. doppelmayeri and M. c. bungei.
The chromosome set of M. c. doppelmayeri does not differ from the chromosome set of M. c. bungei (2n=40, NFa=62) (Fig. 1, f) and consists of 7 pairs of acrocentric chromosomes and 12 pairs of biarmed chromosomes which may be conditionally divided into 7 pairs of meta- and 5 pairs of submetacentric. X-chromosome is a large metacentric, Y - is the least chromosome of the set.
The same chromosome set we described earlier for M. c. camtschatica.
In spite of the considerable morphological divergence of these three subspecies they are ananimous caryologically.
Consequently as it was noticed earlier the morphological resemblance between M. c. doppelmayeri and M. sibirica is convergent. M. sibirica as like the rest eurasian marmots has 38 chromosomes, most of which are biarmed meta- and submetacentric and 4 (5 - M. bobak) acrocentric.
Within limits of Palaearctic marmots M. c. camtschatica forms quite a detailed group. Its karyotipe with a great number of acrocentric chromosomes points out to the adequacy with American marmots.
It may be supposed that M. camtschatica originated from marmots relative to M. caligata (2n=42, NFa=62).
Their karyotypes differ in only one Robertsonian translocation and is the result of at least the second migration of American marmots through the Beringian bridge in early Pleistocene. Nearctic M. broweri (2n=36, NFa=62) probably originated from M. camtschatica (there are differences of their karyotipes in two Robertsonian translocations) and forms the type detached in consequences of the later migration (in the middle, or most likely, in the later Pleistocene) of Asian marmots relative to M. camtschatica through the Beringian land to Alaska.
Now a days has appeared a lot of molecular-genetic methods which provide the possibility to verify our supposition.
We plan to make that in the nearest future. We have already started the comparative studying of the differentially stained chromosomes of marmots. Preliminary study had shown the possibility of intrapopulation variability of localization of nudeolar organizer regions in M. c. bungei. Two exemplars of this species had been studied.
In both animals NOR'S were found on the short shoulders of the first largest pair of meta-submetacentrics (Fig. 1. G. h) and in telomeric regions of two middle by it's size acrocentrics. Except these NORS one specimen has NOR on one of the meta-submetacentric chromosomes of the second pair (Fig. 1, g) (another individual has NORS on the both chromosomes of this pair - Fig. 1, h) and on the long shoulders of chromosomes of the 4th pair (another ones has no NORS on these chromosomes).
In other sciurids, for example, Spermophillus, the localization of NORS is stable inside species, but differs among different species (Korablev, 1984), so it may be a systematic species criteria.
In Marmota as it was shown by our study this feature may be used carefully.
Â
MARMOTA CAMTSCHATICA PALL. - UN ELEMENTO NEARTICO TRA LE MARMOTTE PALEARTICHE
Lyapunova E.A., Boyeskorov G.G., Vorontsov N.N.
N.K. Koltsov Institute of Developmental Biology, the USSR Academy of Sciences, Moscow; Institute of Biology, Sibirian branch of the USSR Academy of Sciences, Yakutsk; USSR Ministry of Natural Resources, Management and Environmental Protection, Moscow
Negli anni '60, prima della scoperta dei metodi di colorazione differenziale, vennero condotti studi cariologici comparativi tra le marmotte oloartiche con i metodi di colorazione routinari. A differenza di gran parte delle marmotte paleartiche, che presentano una notevole omogeneità cariologica (2n=38, NFa=64-66; Vorontsov et al., 1969; Lyapunova et al., 1969; Fernandez-Donoso et al., 1969), eccezion fatta per M. camtschatica (Fig. 1) (2n=40, NFa =62; Lyapunova et al., 1969), le marmotte neartiche mostrano un'ampia divergenza cariologica: M. flaviventris e M. caligata: 2n=42, NFa=62; M. broweri: 2n=36, NFa=62; M. monax: 2n=38, NFa= 64; M. vancouverensis e M. olympus: 2n=40 (Rausch et al., 1965; Hoffmann et al., 1968). Le nette differenze nei livelli di divergenza cariologica tra le marmotte paleartiche e neartiche sono in relazione alla netta allopatria delle specie paleartiche (Fig. 2) ed alla semisimpatria delle marmotte neartiche (Fig. 3). Il carattere di simpatria nella distribuzione e la divergenza nel numero di cromosomi delle specie neartiche riflettono una storia più antica delle forme americane, in confronto alla distribuzione allopatrica ed alla similitudine cromosomica delle specie paleartiche (ad eccezione di M. camtschatica) (Vorontsov et al., 1969; Lyapunova et al., 1969; Vorontsov et al., 1976; Nadler et al., 1976).
L'attuale distribuzione della maggior parte delle specie neartiche é il risultato: della riduzione olocenica degli areali delle specie montane (M. marmota) e di quelle della steppa (M. bobak) o addirittura dell'estinzione (Marmota caucasica); del recupero delle foreste in era post-glaciale (popolazioni ovest-siberiane di M. sibirica e M. camtschactica) e della pressione antropica. In Siberia la distribuzione di M. camtschatica é piuttosto simile a quella di Spermophilus parryi. M. camtschatica occupa le aree montane alpine, mentre S. parryi é presente anche nelle isole steppiche e nella steppa-tundra del nord-est della Siberia.
Entrambe le specie presentano una distribuzione a mosaico, anche se questo tipo di distribuzione é più tipica di M. camtschatica. In questa specie una popolazione può essere separata da un'altra da parecchie centinaia di chilometri.
Le differenze morfologiche tra M. c. camtschatica della penisola Kamtchatka, M. c. bungei della Nord Yakutia e M. c. doppelmayeri della Transbaikalia sono significative (Gromov et al., 1965), cosi come lo sono quelle ecologiche (Bibikov, 1965). In precedenza abbiamo studiato il cariotipo solo delle marmotte della penisola Kamtchatka. à necessario confermare la conspecificità delle differenti popolazioni isolate di sottospecie di M. camtschatica.
Questo lavoro é parte del nostro progetto di revisione della citogenetica delle marmotte olartiche per mezzo delle moderne tecniche di indagine cromosomica.
Materiali e metodi
Sono state analizzate cariologicamente 6 specie di marmotte paleartiche.
- Marmota bobak bobak Muller. Ukraina orientale, distretto di Lugansk, riserva "Streletskaya
step" -1 maschio.
- M. caudata aurea Blanf. 15 esemplari provenienti da tre aree:
2) Pamirs orientali, regione di Murgab, vicino a kishlak Chichekly - 2 maschi e 4 femmine;
3) Thian Shan centrale, catena di Talass, vicino al passo di Besh-Tash - 1 maschio e 2 femmine.
- M. menzbieri zahidovi Petrov. Thien Shan occidentale, catena del Karamin, al limite dell'altipiano di Angren - 1 maschio, 1 femmina.
- M. baibacina centralis Thomas. Thien Shan orientale, Trans-lli Ala-Tau - 1 maschio e 1 femmina.
- M. sibirica sibirica Radde. Transbaikalia, distretto di Chita, nelle vicinanze della stazione ferroviaria di Borzya - 1 maschio.
- M. camtschatica bungei Kastsch. Nord Yakutia: 1) Montagne di Kharaulakh, riva destra del fiume Lena - 1 maschio; 2) Catena del Verkhoyansky, anse superiori del fiume Kele - 2 maschi.
- M. camtschatica doppelmayeri Birula. Buryatia, catena del Barguzin, anse superiori del fiume Urbikan - 1 femmina.
- M. camtschatica camtschatica Pall. Kamtchatka, pendici del vulcano Avacha - 1 femmina.
La preparazione dei cromosomi é stata eseguita secondo la tecnica standard di Ford e Hamerton (1956). I cromosomi dei due maschi di M. camschatica bungei provenienti dalla catena del Verkhoyansly sono stati studiati con la colorazione AgNOR secondo le modifiche di Howell & Black (1980).
Risultati e discussione
Dal momento che i nostri lavori sulle marmotte (Vorontsov et al., 1969; Lyapunova et al., 1969) sono stati pubblicati solo in russo, includeremo in questo lavoro tutti i risultati ed in particolare quelli ottenuti su M. c. doppelmayeri e M. c. bungei. Il set cromosomico di M. c. doppelmayeri non differisce da quello di M. c. bungei (2n=40, NFa=62) (Fig. 1, f) ed é costituito da 7 paia di cromosomi acrocentrici e 12 paia di cromosomi a due braccia, che possono essere suddivisi in 7 paia di metacentrici e 5 paia di submetacentrici, il cromosoma X é grande e metacentrico e il set é completato dal cromosoma Y.
Lo stesso set cromosomico era stato descritto per M. c. camtschatica. Nonostante le notevoli divergenze morfologiche queste tre sottospecie sono cariologicamente omologhe.
Ne consegue, come già dichiarato nel passato, che la somiglianza morfologica tra M. c. doppelmayeri e M. sibirica é di tipo convergente. M. sibirica, come le altre marmotte eurosiatiche, ha 38 cromosomi, la maggior parte dei quali meta - e submetacentrici e 4 (5 in M. bobak) acrocentrici. Tra le marmotte paleartiche, M. c. camtschatica forma un gruppo piuttosto distinto. Il suo cariotipo, con un più alto numero di cromosomi acrocentrici, mette in evidenza la similitudine di questa sottospecie con le marmotte americane. Si può supporre che M. camtschatica derivi da marmotte imparentate con M. caligata (2n=42, NFa=62). I loro cariotipi differiscono per una sola traslocazione Robertsoniana ed é il risultato di una seconda migrazione di marmotte americane attraverso il ponte di Bering, all'inizio del Pleistocene.
La neartica Marmota broweri (2n=36, NFa=62) deriva probabilmente da M. camtschatica (le differenze tra i loro cariotipi sono di due traslocazioni robertsoniane) e costituisce il tipo che si differenziô in conseguenza della successiva migrazione (a metà o, più probabilmente, nella seconda metà del Pleistocene) di marmotte asiatiche imparentate con M. camtschatica verso l'Alaska attraverso la terra di Bering.
Al giorno d'oggi si sono sviluppati numerosi metodi di genetica molecolare che ci consentono di verificare le nostre supposizioni.
Abbiamo in programma di far ciò nell'immediato futuro. Abbiamo già iniziato la studio comparativo di cromosomi di Marmotta colorati con diversi metodi. I risultati preliminari indicano la probabile esistenza di una variabilità , all'interno di una popolazione, della localizzazione delle regioni di organizzazione nucleare (NOR) nella M. c. bungei. Sono stati studiati due esemplari di questa specie. In entrambi gli animali i NOR sono localizzati sui braccio corto del più grande paio di cromosomi meta- e submetacentrici (Fig. 1, g, h) e nella regione telomerica dei due acrocentrici di dimensioni intermedie.
Oltre a questi NOR, un individuo presentava NOR su uno dei cromosomi del seconda paio di meta-submetacentrici (Fig. 1, g) (l'altro individuo possedeva NOR su entrambi tali cromosomi) e sulle braccia lunghe del quarto paio (l'altro individuo non presentava NOR su questi cromosomi).n altri Sciuridi, ad esempio nello Spermophillus sp., la localizzazione dei NOR é costante in ciascuna specie, ma differisce tra le diverse specie (Korablev, 1984) e può essere considerato un carattere sistematico. Il nostro lavoro dimostra che, nel genere Marmota, questa caratteristica deve essere utilizzata con cautela.
Â
REFERENCES
BIBIKOV, D.1. 1967. Mountain marmots of Middle Asia and Kazadhstan. Moscow-Leningrad: 3-703.
COURSER, W., SARGENT, P., BROWNHILL, L.E. and BENIRSCHKE, K. 1963. The somatic chromosomes of the northeastern American woodchuck, Marmota monax. Cytologia (Tokio), 28, 1: 108-111.
FERNANDEZ-DONOSO, R-, FRACCARO, M-, LINDSTEN, J., PORTA, F., SCAPPATICCI, S. and TIEPOLO, L. 1969. The chromosomes of the Alpine marmot Marmota marmota, L- (Rodentia, Sciuridae). Hereditas, 63: 170.
FORD, C.E., HAMERTON, J.L. 1956- A colchicine, hypotonic citrate, squash sequence for mammalian chromosomes. Stain Technol., 31: 247-251.
GROMOV, I.M., BIBIKOV, D.I., KALABUKHOV, N.I. and MEYER, M.N. 1965. The Fauna of the USSR. Mammals. Ground sciuridis. Moscow-Leningrad: 3-446.
GROSSET, G-E-, DINESMAN, L-G- and ZALKIN, V-1. 1965. The ancrent distribution of the steppen marmot. Bull. MOIP, 70, 2: 34-47.
HALL, E.R. and KELSON, K.R. 1959. The mammals of North America. Ronald Press Co., N.Y.: 1-546.
HOFFMANN, R.S. and NADLER, C.F. 1968. Chromosomes and systematics of some North American species of the genus Marmota (Rodentia, Sciuridae). Experientia, 24: 740-742.
HOWELL, W.M. & BLACK, D.A. 1980. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia, 36: 1014-1015.
KAPITONOV, V.I. 1960. Distribution of black-capped marmot (Marmota camtschatica Pall.). Bull. MOIP, 65, 5: 5-15.
KORABLEV, V.P. 1983. Localization of nucleolar organizer regions in karyotype of Palaearctic ground squirrels. VI All-Union symp. on gnavers. Abstr. of papers. Leningrad: 11 7-18.
LYAPUNOVA, E.A., VORONTSOV, N.N. 1969. New data on the chromosome of Eurasian marmots (Marmota, Marmotinae, Sciuridae, Rodentia). The Mammals (Evolution, Karyology, Taxonomy, Fauna). Novosibirsk: 36-40.
NADLER, C.F., HOFFMANN, R.S., VORONTSOV, N.N, and SUKERNIK, R.I. 1976. Evolutionary relationship of some beringian mammals. Beringia in Cenozoic. Vladivostok: 325-336.
PETROV, B.M. 1963. Area of the Menzbier's marmot and fhe western border of the distribution of long-talled marmot in northern Thian Shan. New subspecies of the Menzbier's marmot. Zool. zhurnal, 40, 1: 93-105.
RAUSCH, R. and RAUSCH, V. 1965. Cytogenctic evidence for the specific distribution of an Alaskan marmot, Marmota broweri Hall and Gilmore (Mammalia, Sciuridae). Chromosoma, 16, 5: 618-623.
SAMOSH, B.M. 1958. Causes of the reduction of the area of Marmota bobak in the Ukraine. The problems of land zoo geography.
VAN DER BRINK, F-H- 1958- Die Saugetiere Europas westlich des 30 Langengrades. Hamburg-Berlin, 5:
225.
VERESCHAGIN, N.K. 1959. The Mammals of the Caucasus. Moscow-Leningrad: 3-703-
VORONTSOV, N.N., LYAPUNOVA, E.A. 1976. Genetic aspects of transberingian associations of holarctic mammals. Beringia in Cenozoic. Vladivostok: 337-353.
VORONTSOV, N.N., LYAPUNOVA, E.A., ZAGORUYKO, N.G. 1969. Comparative karyology and evolution of isolating mechanisms in the genus Marmota (Rodentia, Mammalia). Zool. zhurnal, 58, 3: 317-334.
WEHRLI, H. 1935. Die diluvialen Murmeltiere Deutschlands. Palaontol. Z., 17-
Fig. 1. Karyotypes of the palaearctic marmots: a. Marmota bobak, male; b. M. sibirica, male; c. M. baibacina, female; d. M. menzbieri, male; e. M. caudata, female; f. M. camtshatica, male; g. and h: NOR-staining of the chromosomes of two males of M. camtshatica from Yakutia, Verkhoyanye.

Fig. 2: The earlier and the recent distribution of the marmots in Palaearctic. Allopatric character of the distribution, 1 - recent area of M. marmota (acc.to Van den Brink, 1958); - places of finding of the fossil M. marmota (Wehrli, 1935; Grosset et al., 1965) 3 - recreated Quarternary area of M. marmota (Wehrli, 1935; Grosset et al., 1965); 4 - recent area of M. bobak (acc. to diff. authors); 5 - places of findings of the fossil M. bobak (Wehrli, 1935; Samosh, 1958; Grosset et al., 1965; Gromov et al., 1965); - recreated Quarternary area of M. bobak (Wehrli, 1935; Grosset et al., 1965); 7 - traces of findings and supposed area of the fossil Marmota sp. of the Caucasus (Vereschagin, 1959); 8 - recent area of M. baibacina; 9 - the same for M. menzbieri (Petrov, 1963); 10 - the same for M. caudata; 11 - the same for M. hymalayana; 12 - the same for M. sibirica; 13 - the same for M. camtschatica (Kapitonov, 1960); 14 - borders of the recent area of M. camtschatica (Kapitonov, 1960); 15 - place o finding of the fossil M. camtschatica (Grovom et al., 1965); 16 - places of findings of the Quarternary marmots intermediate between M. hymalayana and M. sibirica (Gromov et al., 1965)
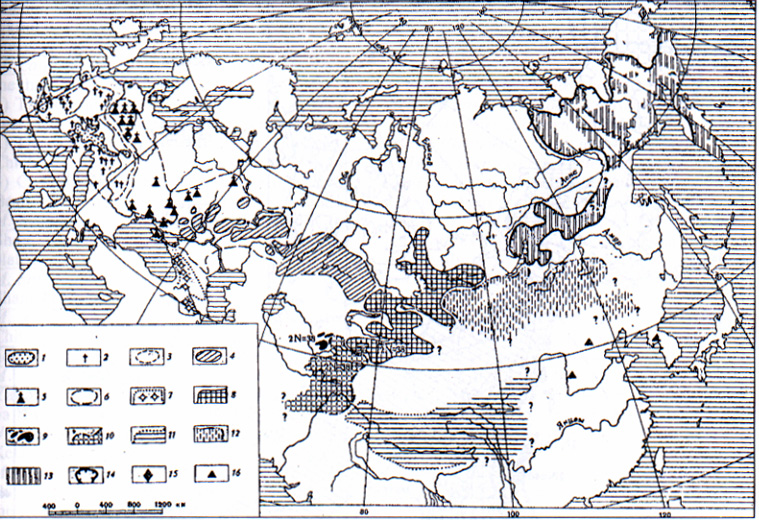
Fig. 3: Semisympatric and sympatric character of the distribution of the American marmots (the areas are composed according to Hall et al., 1959; chromosome numbers - acc.to Rausch et al., 1965 and Couser et al., 1963), 1 - M. camtschatica, 2 - M. broweri, 3 - M. caligata, 4 - M. vancouverensis, 5 - M. olympus, 6 - M. flaviventris, 7 - M. monax
ritorno/back
Tornare index / back contents
Â