Résumé.
Au cours de son cycle saisonnier, la marmotte alpine (Marmota
marmota) présente de remarquables capacités de gestion de ses
réserves lipidiques. La graisse corporelle accumulée durant la
période estivale constitue tout au long de l'hiver l'unique source
d'énergie de cet hibernant. Les modalités de la régulation
hormonale du cycle lipolyse-lipogenèse chez ce mammifère sont
encore de nos jours peu connues. L'objectif de ces recherches est
l'étude de l'évolution saisonnière de la réponse et
de la sensibilité aux hormones des adipocytes prélevés
dans deux dépôts adipeux blancs différents : le tissu
adipeux péritonéal (TAP) et le tissu adipeux sous-cutané
(TASC). Chez les mammifères, la régulation hormonale de ces
processus est assurée principalement par la noradrénaline (NA),
le glucagon, l'insuline et l'ACTH. L'étude de l'action antilipolytique
d'un médiateur local, l'adénosine (ADO), est également
envisagée par le biais d'une enzyme, l'adénosine
désaminase (ADA). Les résultats montrent qu'en période
d'hibernation le métabolisme du tissu adipeux blanc est orienté
vers la mobilisation des substrats. Ainsi, le dépot sous cutané
pourrait soutenir l'approvisionnement basal en substrats
énergétiques ainsi que la mobilisation rapide de ces substrats en
cas de stress, sous l'effet d'une stimulation par l'ACTH. Pour sa part, le
dépôt péritonéal, plus sensible à l'action la
NA, serait le fournisseur principal d'énergie du tissu adipeux brun
(tissu thermogène), lui même fortement solicité au cours
des réveils périodiques. Les résultats obtenus en
présence d'ADA mettent en évidence l'existence d'un
contrôle important de l'activité lipolytique par l'ADO. Les effets
du glucagon et de l'insuline sur l'activité lipolytique des adipocytes
blancs de marmottes fluctuent fortement d'un individu à l'autre et
suivant le dépôt considéré. La sensibilité
des cellules à ces deux hormones reste difficile à
définir. Les cellules ne répondent pas ou peu à l'action
de l'insuline. Cette dernière ne stimule ni l'entrée du glucose
ni son accumulation dans les cellules au cours de la saison d'hibernation. De
même, l'activité de synthèse d'acides gras apparait
fortement réduite durant cette même période.
La quantité de lipides stockés dans les adipocytes résulte du bilan des activités de synthèse et d'hydrolyse des triglycérides. Les vitesses d'hydrolyse (lipolyse) et de synthèse (lipogenèse) des triglycérides peuvent être modifiées par l'environnement hormonal de l'adipocyte in vivo et par sa capacité à répondre aux hormones concernées. Des fluctuations saisonnières de certaines hormones impliquées dans ces processus ont été mises en évidence chez divers hibernants. Les mesures réalisées chez Marmota flaviventris (Tokurama et al. 1991) ont montrées que la glucagonémie ne varie pas significativement au cours du cycle. En revanche, l'insulinémie (et la glycémie) présente un pic automnal coïncidant avec la période de pré-hibernation, caractérisée par une cessation spontanée de la prise alimentaire.
Les modalités de la régulation du cycle lipolyse-lipogenèse, très efficace chez ce mammifère, est encore de nos jours peu connue. L'objectif de nos recherches est d'étudier le cycle annuel de la réponse et de la sensibilité des adipocytes blancs aux hormones impliquées dans cette régulation (noradrénaline (NA), glucagon, ACTH), ainsi que celui d'un médiateur local, l'adénosine (molécule ubiquiste qui est issue en partie de la dégradation de l'ATP, l'AMP cyclique) qui est relarguée par les adipocytes in vivo et in vitro. Cette molécule exerce une action antilipolytique (Shwabe et al. 1976) par l'intermédiaire du récepteur membranaire A1 désormais bien connu (Stiles 1992). Les effets de l'adénosine sont étudiés par l'intermédiaire de l'adénosine désaminase (ADA) qui dégrade la molécule en un composé inactif : l'inosine.
La marmotte est un mammifère herbivore. La majeure partie de l'énergie apportée par les végétaux est de nature glucidique (végétaux pauvres en matières grasses, Florant et al. 1990) Chez les mammifères, le glucose est le principal précurseur de la lipogenèse. L'utilisation du glucose par les adipocytes est limitée par son entrée dans la cellule (James et al. 1989), via deux types de transporteurs dont les caractéristiques sont désormais bien connues.
- Glut 1, qui assure le transport basal du glucose à travers la membrane plasmique des adipocytes à l'état de repos (non stimulés par l'insuline).
- Glut 4, stocké dans des sites intracellulaires et transloqués au niveau de la membrane plasmique sous l'effet d'une stimulation par l'insuline.
L'aspect saisonnier de l'activité lipogénique à partir du glucose peut donc être la conséquence directe de modifications de l'entrée de ce dernier et/ou de son utilisation, processus tous deux fortement dépendants de la sensibilité des adipocytes à l'insuline.
La stimulation des activités lipolytique, lipogénique et du transport membranaire des hexoses est étudiée à partir d'une suspension d'adipocytes isolés, suivant un protocole in vitro décrit par Rodbell (1964). La suspension cellulaire est lavée et reprise dans un tampon Krebs-Ringer dont le pH est de 7,4 , contenant 6 mM de glucose et 5% d'albumine. L'activité lipolytique est mesurée par dosage du glycérol relargué dans le milieu, lors de l'hydrolyse des triglycérides. L'activité lipogénique est mesurée par l'incorporation d'eau tritiée dans les lipides synthétisés au cours de deux heures d'incubation à 37deg.C, sous agitation constante.
L'étude de l'activité lipolytique a été conduite sur deux dépôts graisseux différents : - Le tissu adipeux blanc situé sous le péritoine (dépôt péritonéal), - Le tissu adipeux blanc sous-cutané prélevé au dessus de la région du bas-ventre ou de l'aine (dépôt sous-cutané inguinal). L'étude comparée des deux tissus renseignera sur l'existence éventuelle d'un recrutement préférentiel des triglycérides et des acides gras de l'un ou l'autre de ces deux dépôts.
L'activité de transport du glucose (principal précurseur des acides gras) à travers la membrane adipocytaire est mesurée par l'utilisation d'analogues marqués et non métabolisables du glucose, le 3-O-Méthyl D Glucose (3-O-MG) et le 2 Desoxy-D-Glucose (2-DG). Ces derniers empruntent les mêmes transporteurs que le glucose. Le 3-O-MG traverse la membrane cellulaire via les deux transporteurs Glut 1 et Glut 4 , suivant son gradient de diffusion. L'accumulation de cet hexose s'achève lorsqu'il atteint son équilibre de diffusion. Le 3-O-MG permet de définir l'espace de diffusion cellulaire de l'hexose. Le 2-DG entre dans la cellule selon le même processus mais se concentre au dessus de son équilibre de diffusion (phase de concentration) du fait de sa capacité à subir la première étape de la glycolyse. Il s'accumule dans la cellule sous la forme de 2-DG phosphate (2-DGP). La vitesse de réaction de phosphorylation est plus grande que celle de déphosphorylation. Il se crée dans la cellule une compartimentation biochimique du 2-DGP ce qui ralentit la sortie du 2 DG hors de la cellule (Foley et Gliemann, 1981). L'emploi de cet hexose renseigne non seulement sur la sensibilité des transporteurs à l'insuline mais aussi sur l'efficacité du système de concentration du glucose dans la cellule.
La réponse des adipocytes à la NA et à l'ACTH croît avec la concentration de ces hormones dans le milieu d'incubation. L'effet lipolytique de ces deux hormones est observé chez tous les individus au cours de la saison d'hibernation mais avec des fluctuations interindividuelles importantes. Si l'on se réfère à l'activité lipolytique basale, la réponse maximale est obtenue pour une concentration de 10-5 M de NA et 10-7 M d'ACTH. Les seuils de réponse se situent respectivement autour de 10-7 et 10-9 M, à l'exception des individus 6 et 8, chez qui la NA semble stimuler d'avantage à la concentration 10-8 M que 10-7 M.
Contrairement à ce que l'on peut observer avec la NA et l'ACTH, la réponse des adipocytes au glucagon reste faible chez la marmotte, même aux fortes concentrations d'hormone (sauf pour l'individu 9). Une grande variabilité entre individus est également observée.
L'addition d'ADA dans le milieu d'incubation contribue fortement à l'accroissement de l'activité lipolytique basale Il est intéressant de noter qu'en présence d'ADA, la valeur de l'activité lipolytique stimulée par la NA et le glucagon, quelles que soient les concentrations utilisées, est ramenée à l'activité lipolytique observée en présente d'ADA seule. L'ADA semble restreindre l'expression des propriétés lipolytiques de la NA.
individus sur 6 (individus 6, 8, 9 et 10) dans le cas de l'hormone corticotrope.
En présence d'ADA, les observations précédemment établies évoluent différemment. En effet, les adipocytes du TAP apparaissent d'avantage sensibles à l'ADA entre Octobre et début Décembre. Cette sensibilité à l'ADA tend à décroître dans le TAP et à augmenter dans le TASC au cours de la période comprise entre fin Décembre et Février. Le même phénomène est également observé dans le cas de la réponse à la combinaison NA-ADA.
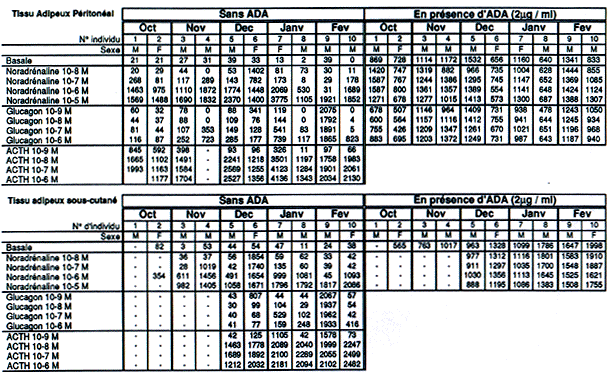
| TA Péritonéal | Oct | Nov | Dec | Jan | Fev | |||||
| Individus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Sexe | M | F | M | M | M | F | F | F | M | M |
| Noradrénaline 10-6 M | 1463 | 975 | 1110 | 1872 | 1774 | 1448 | 2069 | 530 | 31 | 1689 |
| NA 10-6 M + Insuline 10-10 M | - | - | - | 1871 | 2423 | 1251 | 1850 | 1211 | 1703 | 1498 |
| NA 10-6 M + Insuline 10-9 M | 796 | 1030 | 1723 | 2113 | 2301 | 1335 | 2200 | 1123 | 1110 | 1247 |
| NA 10-6 M + Insuline 10-8 M | - | - | 1603 | 2021 | 2337 | 1402 | 2050 | 1060 | 1480 | 1680 |
| Concentration efficace (M) | 10-9 | - | - | SE | - | 10-10 | 10-10 | - | 10-9 | |
| % d'inhibition maximale | 45 | - | - | 0 | - | 14 | 10,5 | - | - | 26 |
Exprimée en nmol de glycérol libéré / 100 ul de cellules / h) d'adipocytes isolés de TAP en présence d'insuline (SE = sans effet)
A partir du mois d'Octobre, trois types de réponses à l'insuline ont été observés. Pour certains individus, les résultats obtenus en présence d'insuline traduisent une évolution inverse de celle habituellement observée chez un mammifère tel que le rat. En effet, pour les individus 3, 5, 8 et 9, l'insuline exerce une franche action lipolytique sur les adipocytes. Aucune raison ne peut actuellement expliquer une telle réponse des cellules adipeuses à cette hormone. Chez l'individu 4, l'insuline n'exerce aucun effet (ni lipolytique, ni antilipolytique), quelle que soit la concentration utilisée. Enfin, les adipocytes prélevés chez les individus 1 et 10 montrent une sensibilité maximale à l'insuline pour une concentration de 10-9 M. L'inhibition de la lipolyse est respectivement de 46 et 26 %. Les adipocytes de l'individu 6 montrent une réponse maximale à l'insuline pour une concentration de 10-8 M. L'inhibition de l'activité lipolytique est alors de 13 %.
| TA sous-cutané inguinal | Oct | Nov | Dec | Jan | Fev | |||||
| Individus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Sexe | M | F | M | M | M | F | F | F | M | M |
| Noradrénaline 10-6 M | - | 354 | 611 | 1456 | 491 | 1654 | 999 | 1081 | 45 | 1093 |
| NA 10-6 M + Insuline 10-10 M | - | - | - | 1120 | 1449 | 1652 | 1176 | 1900 | 1726 | 731 |
| NA 10-6 M + Insuline 10-9 M | - | 315 | 1121 | 1405 | 1269 | 1692 | 1201 | 1986 | 1368 | 349 |
| NA 10-6 M + Insuline 10-8 M | - | - | - | 1391 | 1331 | 1776 | 852 | 380 | 1457 | 642 |
| Concentration efficace (Molaire) | - | 10-9 | - | 10-10 | - | SE | 10-8 | 10-8 | - | 10-9 |
| % d'inhibition maximale | - | 11 | - | 23 | - | 0 | 15 | 64 | - | 68 |
Tableau 3 : Activité lipolytique
Exprimée en nmol de glycérol libéré / 100 ul de cellules / h) d'adipocytes isolés de TASC en présence d'insuline (SE = sans effet)
| TA péritonéal | Oct | Nov | Dec | Janv | Fev | |||||
| Individus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Sexe | M | F | M | M | M | F | F | F | M | M |
| Basale | 26 | 24 | 50 | 71 | 32 | 15 | 30 | 14 | 24 | 21 |
| + ADA (2* ou 4 ug / ml) | 27* | 6* | 57* | 21 | 3 | 3 | 1 | 1 | 6 | 8 |
| + Insuline 10-6 M | 35 | 30 | 66 | 81 | 26 | 13 | 29 | 8 | 31 | 23 |
| + Insuline 10-7 M | 32 | 30 | 101 | 80 | 27 | 13 | 26 | 14 | 34 | 14 |
| + Insuline 10-8 M | 30 | 27 | 101 | 78 | 26 | 13 | 26 | 9 | 28 | 17 |
| + Insuline 10-9 M | 29 | 24 | 95 | 75 | 34 | 11 | 27 | 12 | 29 | 24 |
| Concentration efficace (M) | 10-6 | 10-7 | 10-8 | 10-6 | 10-9 | 10-8 | 10-6 | 10-7 | 10-7 | 10-9 |
| Rapport l. stimulée / l. basale maximal | 1,25 | 2,02 | 1,14 | 1,06 | 0,86 | 0,96 | 1 | 1,41 | 1,14 | |
Exprimée en cpm de 3H incorporés dans les lipides par 100 ul de cellules, au cours d'une heure d'incubation pour 10 millions de cpm initiaux.
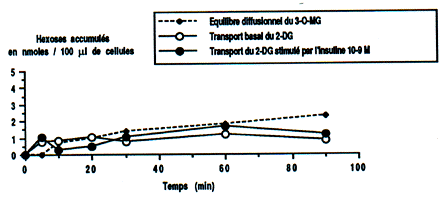
Figure 1 : Étude cinétique du transport des hexoses à travers la membrane adipocytaire.
La vitesse d'entrée et d'accumulation du 3-O-MG et du 2-DG est exprimée en nanomoles pour 100 ul de cellules. l'activité du système concentratif est faible en période de pré-hibernation : le 2-DG ne s'accumule pas au dessus de l'équilibre de diffusion, du 3 -O-MG. On note également une insensibilité à l'insuline du système de transport. La vitesse d'entrée et l'accumulation du 2-DG dans l'adipocytes n'est pas stimulée par l'insuline.
Les résultats obtenus en présence d'ADA mettent en évidence l'existence d'un contrôle important de l'activité lipolytique des adipocytes de marmotte par l'adénosine durant l'hibernation. La levée d'inhibition de la lipolyse basale par l'ADA est forte dans les deux dépôts et un relais dans la sensibilité des tissus à l'adénosine semble prendre place au milieu de la saison d'hibernation. En effet, la sensibilité de l'activité basale du TAP à l'action de l'ADA décroît en milieu d'hibernation alors que celle du TASC augmente à partir de cette période. Il est intéressant de noter qu'en présence d'ADA (donc en absence d'adénosine) dans le milieu d'incubation des cellules, l'effet lipolytique des hormones paraît supprimé et que seul, semble persister celui dû à l'action de l'ADA. Cette observation est contraire à celle faites chez le rat où l'ADA accentue les effets lipolytiques de la NA et du glucagon tout en préservant l'incrément de stimulation provoqué par des doses croissantes d'hormones.
La sensibilité des cellules à l'insuline au cours de l'hibernation reste difficile à définir. L'ensemble des résultats concernant l'activité lipolytique et lipogénique des adipocytes en présence d'insuline tend à montrer que les cellules ne répondent pas ou peu à l'action de cette hormone. L'effet de l'insuline n'est pas reproductible d'un dépôt à l'autre chez les mêmes individus. Les pourcentages d'inhibition maximale mesurés dans le TASC sont supérieurs à ceux du TAP et ne sont pas obtenus pour les mêmes concentrations en hormone.
Des concentrations élevées en insuline ont également peu d'effets sur la stimulation des activités lipogéniques et de transport des hexoses. En milieu de période d'hibernation, les cellules présentent une véritable insensibilité à l'insuline puisque des concentrations de 10-6 et 10-7 M ne suffisent pas à stimuler la mise en réserve du glucose sous forme de lipides. Il en est de même pour le transport des hexoses. Dès novembre l'accumulation du 2-DG dans la cellules ne s'élève pas au dessus de l'équilibre de diffusion. Cette faible accumulation qui n'est pas stimulée par la présence d'insuline dans le milieu d'incubation, montre d'une part que l'hormone ne stimule pas la translocation des transporteurs Glut 4 au niveau de la membrane plasmique de l'adipocytes mais aussi que l'équipement enzymatique nécessaire à la phosphorylation du glucose n'est pas fonctionnel durant cette période du cycle de l'animal.
L'ensemble des résultats semble en accord avec ceux de Tokurama et al, 1990 et Mosfata et al, 1993. Les premiers auteurs ont montré qu'en période de pré-hibernation, l'insulinémie des marmottes à ventre jaune (Marmota flaviventris) est élevée, ce qui serait le reflet d'une diminution de liaison et de la capture de l'insuline par l'ensemble des tissus périphériques dont le tissu adipeux blanc. La raison de cette insulinorésistance n'est pas encore très étudiée chez les hibernants. Les seconds auteurs ont montrés que l'acide gras synthétase ainsi que d'autres enzymes impliquées dans la synthèse et la transformation des acides gras, présentent au cours de l'hibernation une activité minimale dans le tissu adipeux blanc. Les données de la littérature n'indiquent pas si une modification de la vitesse de synthèse est à l'origine de la diminution de l'activité de ces enzymes. De même, aucune recherche concernant un éventuel rythme saisonnier d'une régulation de la transcription des récepteurs à l'insuline n'a été conduite à ce jour.
A suivre...![]()