![]()
Résumé : L'étude comparative du comportement de
fuite de marmottes soumises à différentes pressions anthropiques
a montré une forte accoutumance des marmottes soumises à des
dérangements fréquents. Les distances d'alerte et de fuite des
marmottes peu dérangées sont très nettement
supérieures à celles des marmottes habituées à la
présence de l'homme. Cette différence entre les deux populations
se maintient au cours des mois. Cependant, distances d'alerte et de fuite d'une
population augmentent après l'hibernation. Les marmottes
possèdent donc la capacité de mémoriser les contraintes
afin de s'alimenter pendant l'été, même en présence
de l'homme. Des simulations menées pour tester la notion d'homme
prédateur ou non prédateur n'ont pu aboutir à la
conclusion attendue et aux résultats obtenus en Mongolie sur la marmotte
Bobac. Cette étude devrait être approfondie sur d'autres sites,
où la marmotte est chassée, pour être à même
de conclure sur cet aspect.
Mots-Clés : Marmotte alpine, Marmota marmota,
accoutumance, pression anthropique, prédation, Alpes.
Abstract: Influence of anthropic pressure on the Alpine
marmot
A comparative study about fleeing behaviour of marmots under different
anthropogenic pressure showed a strong familiarization of marmots under
frequent disturbance. Alert and fleeing distance of marmots under fewer
disturbance were strongly greater. The study on several months have shown that
variation between the two populations was always in the same way, even if one
population's median increase after hibernation. So marmots have the capacity to
memorize constraints to forage in summer, when human beings were present.
Simulation conducted for testing notion of human predator or non, have not
ended to the expected conclusion and results obtain in Mongolia on Bobac
marmot. This study has to be extended on other sites, where marmots are hunted
in order to be able to conclude on this aspect.
Key-Words : Alpine marmot, Marmota marmota, familiarization,
anthropic pressure, predation, Alps.
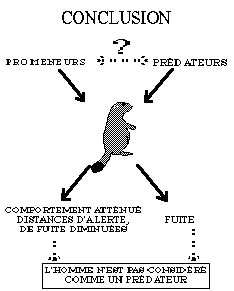
Nous avons aussi voulu savoir si l'homme est considéré par les marmottes comme un prédateur ou non. En effet, les déplacements des promeneurs au cours de la journée empêchent peut-être les renards (Vulpes vulpes) et les aigles (Aquila chrysaetos), qui sont les principaux prédateurs de la marmotte dans les Alpes, d'agir sur les populations. Si l'homme est un prédateur, les marmottes proches du sentier seront plus vigilantes que celles vivant dans un endroit moins fréquenté. Si l'homme n'est pas considéré comme tel, elles peuvent vivre en sécurité tant que la présence des touristes éloigne les prédateurs naturels. Les chasseurs de marmottes de Mongolie sont intéressés par leurs fourrures. Ils veulent pouvoir viser la tête de l'animal afin d'avoir une fourrure intacte. Ils savaient d'expérience qu'elles fuyaient sans donner l'alerte face à un homme, alors que s'il s'agit d'un prédateur, elles se mettent en posture d'alerte, dressées sur les pattes arrière, dans une position favorable à la visée de la tête. Ils avaient donc recours à un costume de loup blanc afin qu'elles donnent l'alarme (Formozov 1994). Ainsi, nous pouvons supposer que les marmottes mongoles ne considèrent pas l'homme en tant que prédateur. Bassano et al. (1992), ont montré que les marmottes alpines ne crient pas à l'approche de l'homme, ce qui confirme l'hypothèse précédente. Pourtant, Nesterova (1994), qui a analysé les différents cris d'alarme chez la marmotte bobac, parle de l'homme en tant que prédateur. Nous avons donc essayé de préciser cette notion de prédateur pour la marmotte alpine, afin d'estimer l'influence --négative ou positive-- de la présence des hommes pendant la saison estivale.
1. Le Vallon de la Sassière, accessible en voiture jusqu'à l'entrée, facile d'accès. Les territoires des groupes étudiés bordent à la fois le sentier principal et un torrent, et leur domaine vital s'étend entre les deux, sur 50 à 100 m.
2. Le Plan du Cheval, plus difficile d'accès à pied, sans chemin. Il est très peu fréquenté, sauf par des groupes de botanistes, et nous permettra de mesurer l'impact de la pression anthropique sur des animaux qui ne sont que rarement en contact avec des promeneurs. Ces données ont été recueillies en juillet 1994 (mois où les promeneurs sont en très grand nombre) et en mai 1995 (mois sans promeneur).
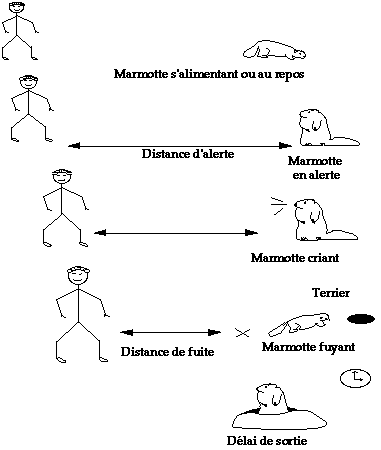
Nous avons effectué quatre mesures (figure 1) :
* Distance d'alerte : l'observateur se déplace en marchant en direction de la marmotte. Il note la position de la marmotte et la distance à laquelle la marmotte réagit à son arrivée : elle lève la tête, le regarde et s'immobilise ou crie, ou bien les deux.
* Distance de fuite : l'observateur continue d'avancer en direction de la marmotte, jusqu'à ce que cette dernière fuie.
* Délai de sortie : lorsque la marmotte pénètre dans un terrier, on déclenche un chronomètre, que l'on arrête dès qu'elle en sort.
En mai, l'un des auteurs s'est déguisé en renard avec une casquette, des vêtements roux et en agitant une queue de renard, afin de représenter un stimulus "supra-normal" de prédateur selon la définition de Tinbergen (1948).
Par contre, ces distances sont significativement différentes entre les deux endroits et sont plus importantes au Plan du Cheval. En juillet, les médianes sont de 91,5 m au Plan du Cheval et de 38,5 m au Vallon (Mann Whitney, z = 4,89 ; p<0,001). En mai, elles sont de 151,5 m et de 78 m respectivement au Plan du Cheval et au Vallon (Mann Whitney, z = 3,48 ; p<0,001). Ces distances sont significativement différentes entre les deux périodes d'études (Mann Whitney, z = 4,22 ; p<0,001 au Plan du Cheval et z = 2,01 ; p<0,001).
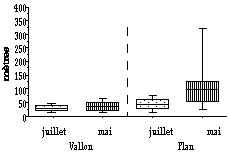
Figure 2. Distances d'alerte Figure 3. Distances de fuite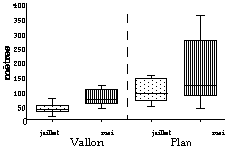
Comme pour les distances d'alerte, les distances de fuite face à la tenue normale et à la tenue de renard (Fig. 3) ne sont pas significativement différentes (Mann Whitney, z = 1,49 ; p>0,05 au Plan du Cheval et z = 0,68 ; p>0,05), et comme précédemment, nous avons cumulé toutes les données.
Les distances de fuite sont significativement plus importantes au Plan du Cheval qu'au Vallon. En juillet, leurs médianes respectives sont 46,5 mètres et 22,5 mètres (Mann Whitney, z = 3,72 ; p<0,001). En mai, elles sont de 96,5 m au Plan du Cheval et de 31,2 m au Vallon (Mann Whitney, z = 4,9 ; p<0,001). Ces distances sont significativement différentes entre les deux mois d'études (Mann Whitney, z = 3,70 ; p<0,001 au Plan du Cheval et z = 2,38 ; p<0,001 au Vallon).
En juillet, sur les 55 observations, seules 5 marmottes ont crié à notre approche (9,1%). En mai, 9 seulement ont crié sur 72 (12,5%).
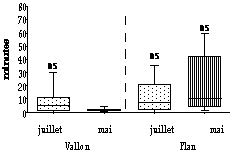
Il n'y a pas de différences entre les délais de latence (Fig. 4) avec les deux tenues (Mann Whitney, z = 0,34 ; p>0,05 au Plan du Cheval et z = 0,55 ; p>0,05 au Vallon). En juillet, les délais de latence ne sont pas significativement différents entre les deux sites d'étude, 7 minutes au Plan et 5,63 minutes au Vallon (Mann Whitney, z = 1,63 ; p>0,005). En mai, ces durées sont significativement différentes entre les deux sites (Mann Whitney, z = 4,7 ; p<0,001). Leurs médianes sont de 10,18 minutes au Plan et de 1,23 minutes au Vallon. Ces durées sont significativement différentes entre les mois de juillet et de mai au Vallon, mais ne le sont pas au Plan du Cheval (Mann Whitney, z = 2,37 ; p<0,05 au Plan du Cheval et z = 0,35 ; p>0,05 au Vallon).
Cette habituation des animaux sauvages à la vue de l'homme a été observée également chez différentes espèces telles que le chamois (Blahout 1977), les oiseaux (Perennou 1990).
Nos résultats sur les délais de latence sont en contradiction avec ceux de la littérature. En effet, Neuhaus et al. (1993) et Gibault (1987), ont montré que les temps de latence des animaux habitués à la présence de l'homme étaient plus courts, alors que dans notre étude, ils ne sont pas différents, sauf en mai. Peut être cette contradiction est-elle liée aux comportements des promeneurs. Au Vallon, ceux qui s'approchent des marmottes sont généralement des photographes qui restent un long moment au bord du terrier principal, pour les surprendre à la sortie. Les marmottes du Vallon se sont familiarisées à cet entêtement et demeurent plus longtemps dans le terrier. Ceci expliquerait qu'en mai, alors qu'il n'y a pas de visiteurs, elles ressortent aussi vite que celles du Plan.
Les animaux semblent capables de distinguer et de reconnaître les différentes espèces qui les entourent et de juger leur danger potentiel. Ainsi, des écureuils terrestres par exemple, peuvent faire la différence entre les prédateurs terrestres et les animaux non dangereux pour eux (Robinson 1980). Les marmottes ne sont plus des gibiers très recherchés en France (Dousset 1984) et sont protégées dans les réserves et les parcs nationaux. Après 20 ans de protection, on peut penser que les marmottes ne craignent plus l'homme en tant que prédateur, ce qui leur permet de s'alimenter en présence d'humains sur le chemin. Nos résultats ne permettent pas de conclure sur cet aspect puisque les marmottes ne sont pas farouches lorsqu'elles voient un humain, mais elles ne le sont pas plus lorsque ce dernier est déguisé en super prédateur. Il serait nécessaire de conduire cette étude sur un lieu ou la marmotte est chassée. Il est également possible que la méthode de chasse pratiquée par les mongols soit une vieille tradition qui s'apparente à un rituel, et que si les chasseurs se décidaient à chasser sans leur déguisement, ils obtiendraient les mêmes résultats.
Les marmottes, comme la plupart des animaux vivant en société, utilisent les cris pour prévenir les autres membres du groupe lorsqu'ils ont repéré un danger. Selon Bassano et al., (1992) les marmottes semblent avoir un cri différent pour les prédateurs terrestres et les prédateurs aériens, ce qui prouve qu'elles sont capables de les reconnaître et de faire passer le message. De la même façon, les singes observés par De Ruiter (1986) se plaquent au sol lorsqu'ils lancent un cri d'alarme d'une prédation aérienne alors qu'une alerte au léopard les fait grimper dans un arbre. Les avis diffèrent cependant sur les cris de la marmotte dirigés vers l'homme. Pour Bassano, les marmottes ne crient pas à l'approche de l'homme, ce qui a été vérifié dans notre travail, puisque seulement 5 marmottes sur 55 et 9 sur 72 ont crié. Nesterova (1994) trouve que les marmottes crient à l'approche de l'homme, alors qu'elles ne criaient pas à l'approche de lapins, chevreuils, et autres animaux non prédateurs. Cependant, Nesterova a fait ses expériences sur la marmotte bobac, qui est encore chassée en ex-URSS. L'homme peut être alors considéré comme un prédateur.
De plus, nous avions suggéré que la présence de l'homme pouvait éventuellement préserver les marmottes de la prédation notamment pendant la journée. Il est vrai que les observations de renard au cours de la journée ont été plus fréquentes au Plan du Cheval, la présence de renard au Vallon, étant surtout observée le soir après 19h00. Compte tenu des capacités cognitives des marmottes, il est probable, qu'elles aient intégré le fait que les hommes n'étaient pas des prédateurs. Les résultats de cette étude ne permettent donc pas de conclure sur l'aspect de la prédation en France.
Les résultats des distances de fuite pourraient être utilisés dans la pratique pour gérer les espaces naturels. Un aménagement des espaces montagnards, se devrait d'étudier auparavant les populations animales locales, incluant les domaines vitaux des espèces présentes, pour faire passer les chemins le plus loin possible des zones sensibles. Cette gestion ne saurait être efficace que si les promeneurs respectent les réglementations en vigueur dans tous les parcs naturels, suivant la loi du 22 juillet 1960, notamment l'interdiction de quitter les chemins.
ALBRECHT L. 1988. Zur Habituation von gemsen (Rupicapra rupicapra L.)
an den Menschen. Diplomarbeit. Zool. Inst. Universität Zürich.
BIDEAU E., GERARD J.F., DESNEUX L. & PICHONI M. 1991. Roe deer introduction
on the mediterranean Peninsula of Sainte Lucie (Aude, France). Relationship
with tourism. In Proceeding of ongulés/ungulates 91, Spitz F.,
Janeau g., Gonzalez g., Aulagnier s. (Eds.), Toulouse, 601-604.
BASSANO B., DURIO P., FACELLO C., PIAZZA R. & TRINCHERO C. 1992.
Preliminary analysis of the temporal and spectral acoustic parameters of
Marmota marmota sound emissions. In 1st Int. Symp. on Alpine Marmot,
Bassano B., Durio P., Gallo Orsi U. et Macchi E. (Eds), Torino, 69-83.
BLAHOUT M. 1977. Niektoré antropické vplyvy na kamziciu zver v
tatranskom narodnom parku. Zbornik prac o tatranskom narodnom parko, 19:
127-168.
DE RUITTER J.R. 1986. The influence of group size on predator scanning and
foraging behavior of wedgecapped capuchin monkeys (Cebus olivaceus).
Behaviour, 98: 240-258.
DOUSSET L. 1996. Une transformation structurale : évolution du rapport
homme-marmotte (Marmota marmota) dans les alpes françaises. / A
structural transformation: the evolution of the relationship between people and
marmot (Marmota marmota) in the French Alps. In Biodiversité
chez les marmottes/Biodiversity in Marmots, Le Berre M., Ramousse R. &
Le guelte L., Eds, International Network on Marmots, 29-36.
FORMOZOV N.A., YENDUKIN A. YU. & BIBIKOV D.I. 1996. Coadaptations des
marmottes (Marmota sibirica) et des chasseurs de Mongolie. /
Coadaptations of marmots (Marmota sibirica) and mongolian hunters. In
Biodiversité chez les marmottes/Biodiversity in Marmots, Le Berre
M., Ramousse R. & Le guelte L., Eds, International Network on Marmots,
37-42.
GAILLARD J.M., VASSANT J. & KLEIN F. 1987. Quelques
caractéristiques de la dynamique des populations de sangliers (sus
scrofa scrofa) en milieu chassé. Gibier Faune Sauvage, 4:
31-47.
GIBAULT C. 1994. Plasticité du comportement et régime
alimentaire de la marmotte alpine (Marmota marmota) sous différentes
pressions anthropiques. Rapport de DEA "Sciences du comportement et
neurosciences cognitives", Toulouse.
GIBAULT C., R. RAMOUSSE & M. LE BERRE 1996. Effets de la pression
anthropique sur le comportement alimentaire de la marmotte alpine. / Hiking
influence on feeding behaviour in Alpine Marmot. In Biodiversité chez
les marmottes/Biodiversity in Marmots, Le Berre M., Ramousse R. & Le
guelte L., Eds, International Network on Marmots, 233-234.
NEUHAUS P., MAININI B. & INGOLD P. 1993. Human impact on marmot behaviour.
In 1st Int. Symp. on Alpine Marmot. Bassano B., Durio P., Gallo Orsi U.
et Macchi E. (Eds). Torino, 165-169.
NESTOROVA N.L., 1994. Comportement d'alarme et réponse au cri d'alarme
en fonction de l'âge chez les marmottes bobacs. /Age-dependent alarm
behavior and response to alarm call in bobac marmots (Marmota bobac
mull). In Biodiversité chez les marmottes/Biodiversity in
Marmots, Le Berre M., Ramousse R. & Le guelte L., Eds, International
Network on Marmots, 181-186
PERENNOU C. 1990. Peuplements d'oiseaux aquatiques en milieu
anthropisé : un exemple. Les plaines de la côte de Coromandel
(Inde du Sud-Est). Thèse de doctorat de l'Université Claude
Bernard- Lyon 1.
ROBINSON S.R. 1980. Antipredator behavior and predator recognition in belding
ground squirrels. Anim. Behav., 28: 840-852.
TINBERGEN N., 1948. Social releasers and the experimental method required for
their study. Wilson Bull., 60, 6-51.
FRANCESCHINA-ZIMMERLI R. & INGOLD P. 1994. Comportement des marmotes
alpine, Marmota m. marmota, sous différentes pressions de
randonneurs. /The behavior of alpine marmots (Marmota m.marmota) under
different hiking pressure. In Biodiversité chez les
marmottes/Biodiversity in Marmots, Le Berre M., Ramousse R. & Le Guelte
L., Eds, International Network on Marmots, 73-74.
![]()