(MARMOTA MARMOTA)
![]()
Résumé : Afin d'estimer l'âge individuel de
Marmota marmota nous avons utilisé une technique fondée
sur des mesures biométriques effectuées sur le vivant. Deux
populations, dont l'âge des individus était connu, ont
été choisies comme référence. Pour chacune, nous
avons pris en compte les mesures biométriques, l'âge et le sexe
des marmottes lors de session de captures (1993 et 1994). Les individus ont
été rangés dans 4 classes d'âge : marmottons,
marmottes de un-an, marmottes de 2 ans et adultes. La discrimination de ces
classes d'âge à partir de l'ensemble des mesures
biométriques met en évidence que la masse corporelle et la
longueur totale du corps sont de bonnes variables. Mais, il n'est possible de
discriminer que 3 classes d'âge : les marmottons, les marmottes de 1 an
et les marmottes de 2 ans et plus.
Mots-clés : Marmota
marmota, Biométrie, âge.
Abstract: Contribution of biometry to age determination of Alpine
marmot (Marmota marmota).
In order to evaluate the age of Marmota marmota, we used a technic
based on biometrical measures on living animals. Two populations of animals
with known ages were used as reference. For each of them we took into account
the biometrical measures, age and sex during trapping sessions (1993 and 1994).
The animals were divided in four age classes: cubs, yearlings, two-year-old and
adults. Taking all the measures into account showed that body mass and overall
length of the body are good measures of discrimination of the age classes. But
only three age classes appeared clearly, cubs, yearlings and two year-old or
more than two year-old individuals. Key-Words: Marmota marmota,
biometry, age.
Certaines de ces méthodes ont été appliquées aux marmottes. Par exemple, chez Marmota monax, Davis (1964) utilisant les caractéristiques des organes reproducteurs, du pelage et des dents, le poids du cristallin, la masse corporelle et la longueur du corps, conclut qu'aucun de ces facteurs n'est discriminant de l'âge. De même, chez Marmota flaviventris, Armitage et al. (1976) montrent l'imprécision de la masse corporelle pour l'estimation de l'âge. Par contre, chez Marmota monax Nuckle & Bergeron (1983) ont mis en évidence, à l'aide de la méthode du poids du cristallin, trois classes d'âge : marmottons, un an et plus de 2 ans. Seulement, cette méthode s'applique à des animaux morts, or le gestionnaire de faune a besoin de méthodes utilisables sur le vivant. C'est pourquoi, à partir de deux populations de marmottes alpines d'âge connu, nous proposons une technique d'estimation de l'âge qui utilise des mesures biométriques effectuées sur le vivant.
Chaque animal capturé est sexé (distance entre l'anus et l'orifice génital). L'âge des animaux est déterminé par le suivi des populations, année après année et par la prise en compte de la masse corporelle de l'animal. La reconnaissance des marmottons et des un-an est aisée en début de saison. Par contre, la détermination de l'âge, au delà de 2 ans, est difficile voire impossible, sans un suivi des individus. Les marmottes sont regroupées en 4 classes d'âge : - classe 1 : les marmottons, - classe 2 : les marmottes de un an, classe 3 : les marmottes de 2 ans, classe 4 : les marmottes de 3 ans et plus.
Enfin, une série de mesures est réalisée sur chaque animal : 1- masse corporelle (grammes) ; 2 - longueur de la tête et du corps (cm) ; 3 et 4- longueur des pattes antérieures LPA et postérieures LPP (mm) ; 5 - distance ano-génitale DAG (mm) ; 6- tour du cou TC (mm) ; 7- largeur de la tête LT (mm) ; 8- la longueur des maxillaires LM (mm) ; 9 et 10- la longueur des incisives inférieures LII et supérieures LIS (mm). Pour l'analyse, nous avons retenu 194 individus pour le site de la Sassière (101 femelles et 93 mâles) et 123 individus pour le site de Prapic (62 femelles et 61 mâles). Sur les deux sites, les mesures morphométriques ont été réalisées par plusieurs personnes.
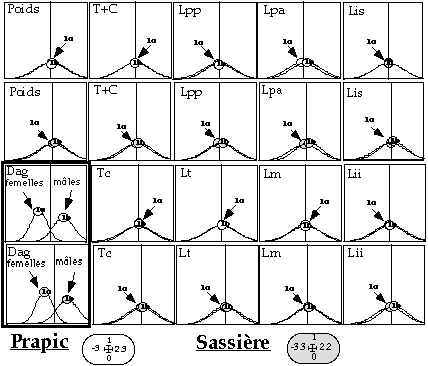
La présence éventuelle d'un dimorphisme sexuel chez la marmotte a été recherchée (test de Mann-Whitney entre mâles et femelles). Puis, le pouvoir discriminant de chaque variable biométrique a été testé par un test de Kruskal-Wallis entre classes d'âge. Enfin, une analyse discriminante sur l'ensemble des valeurs biométriques d'un individu, permet d'estimer quelles sont les variables biométriques les plus intéressantes dans la détermination de l'âge individuel.
| Prapic | Sassière | |||
| Z corrigé | p | Z corrigé | p | |
| Poids | -0,45 | 0,6541 | -0,77 | 0,4425 |
| T+C | -0,08 | 0,9334 | -1,58 | 0,1133 |
| Lpp | -0,44 | 0,6632 | -2,04 | 0,041 |
| Lpa | -1,43 | 0,1515 | -1,95 | 0,0518 |
| Dag | -8,78 | 0,0001 | -10,76 | 0,0001 |
|---|---|---|---|---|
| Tc | -0,27 | 0,7863 | -1,22 | 0,2224 |
| Lt | -0,2 | 0,8454 | -1,11 | 0,2672 |
| Lm | -0,19 | 0,8534 | -0,68 | 0,4958 |
| Lis | -0,28 | 0,7813 | -1,67 | 0,0948 |
| Lii | -0,83 | 0,4041 | -0,99 | 0,321 |
| Prapic | Sassières | |||
| H corrigé | p | H corrigé | p | |
| Poids | 94,4 | 0,0001 | 153,4 | 0,0001 |
| T+C | 101,5 | 0,0001 | 164,1 | 0,0001 |
| Lpp | 88,8 | 0,0001 | 138,3 | 0,0001 |
| Lpa | 54,8 | 0,0001 | 125,2 | 0,0001 |
| Dag | 19,6 | 0,0002 | 49,9 | 0,0001 |
| Tc | 99 | 0,0001 | 150,3 | 0,0001 |
| Lt | 77,9 | 0,0001 | 146,3 | 0,0001 |
| Lm | 88,6 | 0,0001 | 156,7 | 0,0001 |
| Lis | 93,8 | 0,0001 | 161,2 | 0,0001 |
| Lii | 89,8 | 0,0001 | 159,2 | 0,0001 |
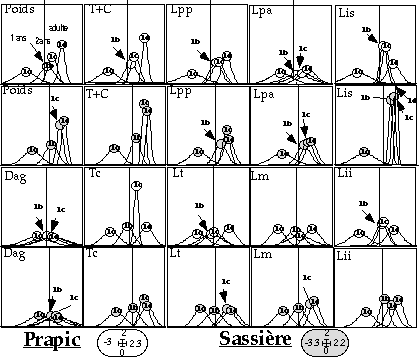
La mesure de la distance ano-génitale apparaît peu discriminante de l'âge. Notons que les variances entre classes ne sont pas identiques. La variance observée pour la classe des marmottons est supérieure, en grande partie à cause de la croissance. Sinon, l'inégalité des variances entre les autres classes s'expliquerait par des différences d'effectif, notamment pour le site de Prapic où, la classe 3 (2 ans) n'est représentée que par 8 individus contre 30, 45 et 40 marmottes pour les classes 1, 2 et 4 respectivement. L'étude univariée ne nous permet donc pas de connaître quelles sont les variables les plus discriminantes. C'est à partir des analyses discriminantes qu'il est possible de faire ressortir une structure globale aux deux sites étudiés :
- trois classes d'âge peuvent être réellement distinguées, les marmottons, les marmottes de un an, et celles de plus de deux ans (fig. 3 et 4). Sachant que la maturité sexuelle est atteinte à l'âge de 2 ans, ce résultat serait justifié. La marmotte est supposée avoir alors une taille d'adulte et une croissance nulle.
- la masse corporelle et la longueur totale (T+C) sont les variables les plus discriminantes pour les deux sites (figure 5). De même, la distance ano-génitale (Dag) et la longueur des pattes antérieures sont les variables les moins discriminantes.
- la présence d'une évolution le long de l'axe F1 des marmottons aux adultes.
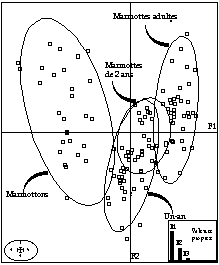
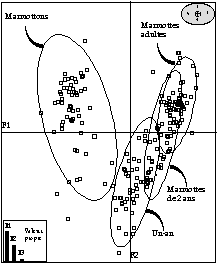
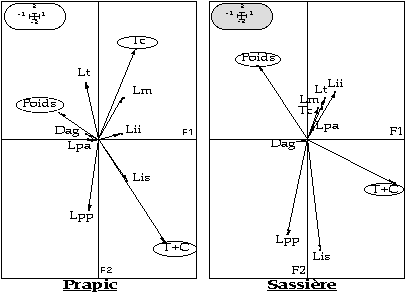
L'intérêt d'un tel travail est de déterminer à partir de mesures sur le vivant, la structure d'âge d'une population inconnue. L'analyse discriminante serait une technique pour assigner efficacement un spécimen correspondant à une catégorie d'âge. Cette démarche paraît d'un intérêt certain pour les gestionnaires de populations d'animaux sauvages. Afin d'affiner la technique, trois directions seraient à développer sur ce sujet :
1- Réaliser de nouvelles analyses sur de nouveaux jeux de données : ajouter les années 1995 et 1996 afin de vérifier la similarité des résultats.
2- Etablir une population de référence pour un site donné à partir d'un échantillon de crânes recueillis.
3- Etudier l'effet des conditions du milieu sur la "morphologie" des marmottes.