Université Joseph Fourier, BP 53,
F-38041 Grenoble Cedex 9, FRANCE
(2) Laboratoire de Biologie des Populations d'Altitude, URA 1946,
Université Claude Bernard Lyon 1
43 boulevard du 11 Novembre 1918, F-69622 Villeurbanne, FRANCE
![]()
![]()
Résumé : Un cas d'acceptation d'une femelle gravide et
d'adoption de ses jeunes par le mâle dominant d'un groupe familial de
marmottes alpines (Marmota marmota) a été confirmé
par l'étude conjointe de l'analyse du polymorphisme de 6
séquences microsatellites et d'observations comportementales. Ces
premiers résultats semblent indiquer que le système de
reproduction de la marmotte alpine est plus complexe qu'on ne le croyait, que
la polygynie n'est pas exclue et que des femelles adultes peuvent joindre des
groupes voisins. Ce transfert aurait été interprété
comme une paternité extra-couple si des données de terrain
très complètes n'avaient été disponibles.
Mots clés : Marmota marmota, paternité,
système de reproduction, poil, PCR, microsatellites
Abstract : Immigration of a pregnant female and adoption of
its offspring in a family group of Alpine marmot. Genetic and behavioural
data.
The acceptance of a pregnant female and the adoption of its offspring by the
dominant male of a family group of Alpine marmot (Marmota marmota) was revealed
both by microsatellite polymorphism analysis and behavioural studies. These
preliminary results show that the mating system of the Alpine marmot is more
intricate than believed. Polygyny cannot be excluded and adult females can
aggregate neighbouring groups. Such aggregation would have been misinterpreted
as an extra-pair paternity if very precise field data had not been
available.
Key-words : Marmota marmota, paternity, mating system, hair, PCR,
microsatellites.
La marmotte alpine fait partie d'un genre monophylétique présentant une complexité dans la structure sociale qui en fait un modèle particulièrement intéressant pour l'étude de l'évolution de la socialité chez les mammifères. Marmota marmota semble monogame mais des cas de paternités extra-couples ont été suggérés (Arnold 1990a). Les accouplements se produisant généralement à l'intérieur des terriers, il est difficile d'identifier le père de chaque jeune. Rassmann et al. (1994) ont révélé, par les minisatellites, un faible degré de polymorphisme dans les populations étudiées. Preleutner et Pinsker (1993) ont également révélé, par les allozymes, une faible variabilité génétique dans les populations autrichiennes. Dès lors, l'analyse des paternités nécessite un marqueur génétique plus polymorphe, les microsatellites qui sont des courtes séquences répétées de 1 à 4 paires de bases, dispersées de façon régulière et au hasard dans tout le génome des Eucaryotes, et montrant un fort taux de mutation. Nous présentons ici les premiers résultats d'une étude du système de reproduction de la marmotte alpine. Des données génétiques et comportementales ont permis de mettre en évidence le cas de l'acceptation d'une femelle gravide par le mâle d'un groupe voisin.
Les analyses hormonales ont permis de mettre en évidence que les trois femelles FdB, FsB et FdA étaient pleines.
Les lignes 1 et 2 correspondent aux mâles adultes MdA et MsA, les lignes 3 et 4 corresondent aux femelles adultes FdA et FsB, les lignes 5, 6, 7 et 8 correspondent aux jeunes du groupe A. La ligne 9 correspond au mâle adulte du groupe B, les lignes 10, 11 et 12 correspondent aux mâles adultes du groupe C et les lignes 13 et 14 correspondent aux mâles adultes du groupe D. A, B, C, D et E sont les noms des allèles des groupes A, B, C et D.
PCR products on polyacrylamide colloid denaturing the SS-Bibl1 locus in the A family group of the Grande Sassière and in the males of the neighbouring groups.
Lines 1 and 2 correspond to the adult males MdA and MsA, Lines 3 and 4 to the adult females FdA and FsB, lines 5, 6 7, 8 to the offspring of the A group. Line 9 correspond to the adult male of the B group, lines 10, 11, 12 to the adult males of the C group and lines 13 and 14 to the adult males of the D group. A, B, C, D point to tha allels of the A, B, C, D groups .
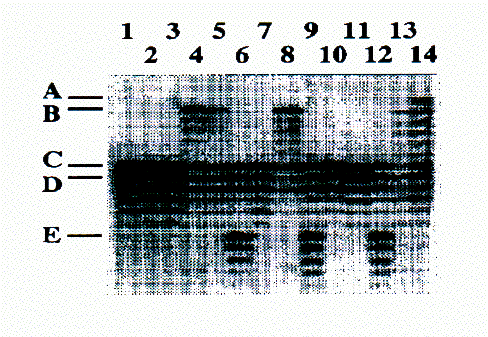
Tableau 1. Exclusions de paternité et variation allélique pour les individus des groupes familiaux A et B et pour les mâles adultes des groupes voisins (C et D). Les allèles des jeunes, en caractère gras, sont les allèles incompatibles avec ceux des mâles adultes du groupe A.
Paternity exclusionsand allel variationin individuals of the family groups A and B, and in males of the neighbour groups (C and D). Incompatible allels of offspring with those of the adult males of A group are in bold characters.
| Référence | Sexe | Statut dans le groupe | Locus SSBibl1 | Locus SSBibl4 | Locus SSBibl18 | Locus SSBibl20 | Locus SSBibl25 | Locus SSBibl31 |
| Groupe A | ||||||||
| MdA | Mâle | Adulte | C/C | F/F | K/K | N/O | Q/R | U/V |
| MsA | Mâle | Subadulte | C/C | F/G | H/K | L/N | Q/Q | U/U |
| FdA | Femlle | Adulte | C/C | F/G | J/K | M/O | R/R | U/V |
| FSB | Femelle | Adulte | B/C | F/G | I/K | N/O | P/R | U/U |
| S8031 | Femelle | Un-an | C/C | F/F | K/K | N/O | Q/R | U/V |
| S8035 | Femelle | Un-an | C/C | F/F | K/K | N/O | Q/R | T/U |
| T7643 | Femelle | Un-an | C/C | F/G | J/K | M/N | P/R | T/V |
| T7928 | Mâle | Un-an | C/C | F/F | J/K | M/N | Q/R | U/V |
| T7932 | Mâle | Un-an | C/C | F/F | J/K | N/O | Q/R | U/V |
| T7545 | Femelle | Marmotton | B/C | F/G | I/K | M/O | Q/R | U/V |
| T7268 | Femelle | Marmotton | C/E | F/F | I/I | N/O | Q/R | U/V |
| T7249 | Mâle | Marmotton | C/C | F/G. | I/K | M/O | P/R | U/V |
| T7213 | Mâle | Marmotton | B/C | F/F | I/I | N/O | P/R | U/V |