Spectral analysis shows that these whistles are quite different from those recorded with other models; Fme is highest (P<0.01) and also a.d. (P<0.01) in fact second
harmonic is frequently absent (fig. 1). Geometrical models seem to represent a kind of stimulus quite different from others: there are no differences changing rectangular dimension nor it is possible to reproduce the effects due to other models of similar dimension. Marmots show more concern for predators' shapes than for non predators' ones (difference as number of whistles). In the case of predator raptors models, first call seems correlated with danger kind or degree, while following vocalizations are used to manteining high attention and alert in the colony until danger is ended. The model's silhouette greatly influences acoustic antipredatory alert behaviour and surely more than model's dimension. Although models and real birds represent two different perceptive realities, it's most probable that differences seen testing the animals' reactions to artificial shapes are just due to the simple aspects (as dimension or silhouette) that these stimuli show.
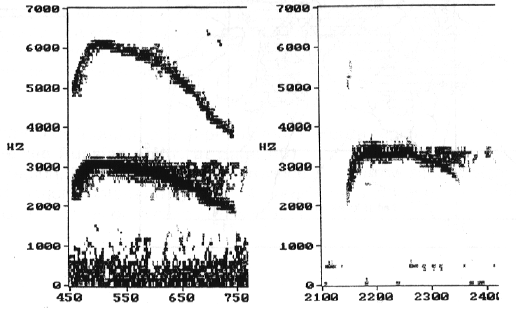
Fig. 1. Left sonogram (band width 43 Hz) shows a whistle with a high amplitude of second harmonic (most typical of the first call). On the right a call whose second harmonic is almost absent (most typical of secondly whistles and of calls related to geometrical shapes).
Back to Abstracts contents