Marie-Pierre Callait
Laboratoire dﺣ۸partemental d'analyses vﺣ۸tﺣ۸rinaires, Chambﺣ۸ry
Rﺣ۸sumﺣ۸ : La structure des systﺣ۷mes hﺣﺑtes-parasites et les diffﺣ۸rents niveaux de distributions des parasites dans la population-hﺣﺑte pendant la fin de la pﺣ۸riode d’activitﺣ۸ de la marmotte alpine, phase maximale de l’intensitﺣ۸ parasitaire ont ﺣ۸tﺣ۸ examinﺣ۸s. Les helminthes ont ﺣ۸tﺣ۸ recherchﺣ۸s chez vingt-deux marmottes provenant du mﺣ۹me site. Des trois principales espﺣ۷cesﺡ parasites observﺣ۸es, Ctenotaenia marmotae prﺣ۸sente la prﺣ۸valence, l’abondance et l’intensitﺣ۸ moyenne les plus ﺣ۸levﺣ۸es, Citellina alpina, les plus faibles, et Ascaris laevis a des valeurs intermﺣ۸diaires. Par contre, si ces trois espﺣ۷ces prﺣ۸sentent un ﺣ۸tat d’agrﺣ۸gation , celui-ci est trﺣ۷s ﺣ۸levﺣ۸ chez Citellina et diminue chez Ascaris puis Ctenotaenia. Les stratﺣ۸gies d’infestation, de transmission et de survie sont discutﺣ۸es en tenant compte des caractﺣ۸ristiques des espﺣ۷ces-parasites et de l’espﺣ۷ce-hﺣﺑte. Elles traduisent les phﺣ۸nomﺣ۷nes de co-ﺣ۸volution trﺣ۷s poussﺣ۸s qui unissent parasites et hﺣﺑte. Mots clﺣ۸s : Marmota marmota, systﺣ۷mes hﺣﺑtes-parasites, helminthes, dynamique parasitaire, Savoie.
Abstract: Distribution of the parasite infrapopulations in the Alpine marmot.
Helminth-marmot systems and the distribution of the parasites in an host population were studied at the end of summer, phase of maximum parasite intensity. Parasitic surveys of twenty two marmotsprincipales at the end of summer, phase of maximum parasite intensity. Parasitic surveCtenotaenia marmotaeysCitellina alpina, Ascaris laevis of twenty two marmotse and average intensity were highest in Ctenotaenia , lowest in Citellina and intermediate in Ascaris. But all the species had high level organisation with over-dispersed patterns of distribution, decreasing from. Citellina to Ctenotaenia. Infestation, transmission and survival strategies, depending on the characteristics parasite-species and host-species, were discussed in relation with co-evolution phenomenon.Key-words : Marmota marmota, host-parasit systems, helminths, parasitic dynamics, Savoy, France.
Pour chaque espﺣ۷ce parasite, la description de la distribution individus-parasites dans les individus-hﺣﺑtes est essentielle dans l’ﺣ۸tude des phﺣ۸nomﺣ۷nes densitﺣ۸-dﺣ۸pendants qui ont lieu au niveau de la population parasite (Poulin 1993). La distribution des parasites parmi les hﺣﺑtes est utilisﺣ۸e comme base de l’ﺣ۸valuation quantitative de la nature du parasitisme (Crofton 1971a). Il est donc indispensable de pouvoir distinguer les distributions observﺣ۸es et de les relier ﺣ des distributions thﺣ۸oriques connues, qui pourront servir de modﺣ۷le pour comprendre les mﺣ۸canismes qui les dirigent. La distribution de trois espﺣ۷ces d’helminthes a ﺣ۸tﺣ۸ ﺣ۸tudiﺣ۸e chez un ﺣ۸chantillon de marmottes alpines sur le site de Bonneval-sur-Arc (Savoie) pendant la saison de chasse.
1. Objectifs et mﺣ۸thodes
Nous disposons des rﺣ۸sultats de 22 bilans parasitaires, issus de l’examen, soit de cadavres entiers, soit uniquement de viscﺣ۷res. Seuls les helminthes ont ﺣ۸tﺣ۸ recherchﺣ۸s dans cette partie de l’ﺣ۸tude. L’ﺣ۸chantillon provient d’animaux tuﺣ۸s ﺣ la chasse sur le site de Bonneval-sur-Arc, au cours des mois de septembre et d’octobre 1996 et 1997. Leur statut social au sein de la colonie n’est pas connu.
Les index ﺣ۸pidﺣ۸miologiques (prﺣ۸valence, abondance, intensitﺣ۸ moyenne) sont calculﺣ۸s pour chaque infrapopulation parasite ﺣ partir des rﺣ۸sultats de l’ﺣ۸chantillon entier.
Le test de Normalitﺣ۸ de Kolgomorov-Smirnov (probabilitﺣ۸s de Lilliefors) est appliquﺣ۸ aux donnﺣ۸es observﺣ۸es pour dﺣ۸terminer la nature des tests statistiques ﺣ utiliser par la suite (paramﺣ۸triques ou non-paramﺣ۸triques). Le logiciel utilisﺣ۸ pour les calculs statistiques est STATISTICA version 5 (StatSoft).
Il est possible de caractﺣ۸riser le mode de dispersion des organismes par des distributions statistiques pour des variables biologiques discontinues (Combes 1995, Frontier & Pichon-Viale 1998)ﺡ : elles sont agrﺣ۸gﺣ۸es ("concentration" des parasites dans certains individus-hﺣﺑtes, au dﺣ۸triment d’autres), "ﺡ poissonniennesﺡ " (rﺣ۸partition au hasard) ou rﺣ۸guliﺣ۷res (rﺣ۸partition identique chez tous les individus-hﺣﺑtes). L’agrﺣ۸gation est trﺣ۷s rﺣ۸pandue en ﺣ۸cologie parasitaire (Combes 1995, Crofton 1971a et b).
La distribution de frﺣ۸quences du nombre de parasites par hﺣﺑte illustre quelle proportion de la population-hﺣﺑte porte un nombre donnﺣ۸ de parasites. De plus, plusieurs indices peuvent ﺣ۹tre calculﺣ۸s pour caractﺣ۸riser la tendance d’une distribution observﺣ۸e et en particulier le niveau d’agrﺣ۸gation.
Si R est voisin de 1, la distribution est "ﺡ poissonnienneﺡ " (variance ﺣ۸gale ﺣ la moyenne)ﺡ ; s’il est infﺣ۸rieur ﺣ 1, elle est rﺣ۸guliﺣ۷reﺡ ; s’il est supﺣ۸rieur ﺣ 1, elle est agrﺣ۸gﺣ۸e. Elle est d’autant plus agrﺣ۸gﺣ۸e que R est ﺣ۸levﺣ۸. Il permet de plus de comparer des ﺣ۸chantillons de prﺣ۸valence et d’abondance diffﺣ۸rentes.
Nous utiliserons l’estimateur du maximum de vraisemblance k3, car les conditions nﺣ۸cessaires au calcul des deux autres estimateurs ne sont remplies par aucune des distributions (Scherrer 1984). Cet estimateur k3 correspond ﺣ une ﺣ۸quation itﺣ۸rative et ne nﺣ۸cessite pas de condition particuliﺣ۷reﺡ :
![]() avec
avec ![]() = somme des frﺣ۸quences observﺣ۸es des classes supﺣ۸rieures ﺣ yi.
= somme des frﺣ۸quences observﺣ۸es des classes supﺣ۸rieures ﺣ yi.
k est une estimation indirecte de l’agrﺣ۸gation, car il est issu d’une distribution binomiale nﺣ۸gative thﺣ۸orique.
Le calcul de D, aprﺣ۷s simplification, est le suivantﺡ :

oﺣﺗ N = le nombre d’hﺣﺑtes dans l’ﺣ۸chantillonﺡ ; xj = le nombre de parasites dans l’hﺣﺑte j (les hﺣﺑtes ﺣ۸tant classﺣ۸s par ordre croissant de charge parasitaire)ﺡ ; ![]() = le nombre moyen de parasites par hﺣﺑte.
= le nombre moyen de parasites par hﺣﺑte.
L’index a pour valeur minimale 0, quand la courbe observﺣ۸e correspond ﺣ la ligne droite hypothﺣ۸tique, et pour valeur maximale 1, quand tous les parasites sont portﺣ۸s par un seul hﺣﺑte (agrﺣ۸gation maximale). Dobson (1986) souligne qu’il faut toujours associer l’index D et l’histogramme de Lorenz correspondant. En effet, une valeur de l’index peut reprﺣ۸senter deux types de distributionsﺡ : soit beaucoup d’hﺣﺑtes portent peu de parasites et beaucoup de parasites sont portﺣ۸s par peu d’hﺣﺑtes (cas le plus frﺣ۸quent), soit beaucoup d’hﺣﺑtes portent beaucoup de parasites et peu de parasites sont portﺣ۸s par peu d’hﺣﺑtes. La courbe de Lorenz correspondant au second cas est symﺣ۸trique de la premiﺣ۷re mais au-dessus de la ligne droite de distribution idﺣ۸ale.
La prﺣ۸cision de l’estimateur de l’agrﺣ۸gation augmente avec la taille de l’ﺣ۸chantillon (Poulin 1996). Le degrﺣ۸ d’agrﺣ۸gation ne dﺣ۸pend pas de la faﺣ۶on dont les hﺣﺑtes sont examinﺣ۸s, mais les hﺣﺑtes fortement infestﺣ۸s qui caractﺣ۸risent l’agrﺣ۸gation sont rares et n’apparaissent que dans les grands ﺣ۸chantillons. Toutes les mesures de l’agrﺣ۸gation tendent ﺣ sous-estimer l’agrﺣ۸gation rﺣ۸elle du systﺣ۷me hﺣﺑte-parasite ﺣ۸tudiﺣ۸ lorsque l’ﺣ۸chantillon est rﺣ۸duit.
Nous chercherons ﺣ expliquer les patrons de distribution des parasites en fonction des particularitﺣ۸s des espﺣ۷ces-parasites (mode de transmission, prolificitﺣ۸, rﺣ۸sistance et distribution des formes de dissﺣ۸mination dans le milieu extﺣ۸rieur), des caractﺣ۸ristiques des espﺣ۷ces-hﺣﺑtes (individusﺡ : sexe, ﺣ۱ge et statut socialﺡ ; populationsﺡ : organisation sociale et spatiale, nature et intensitﺣ۸ des contacts, homogﺣ۸nﺣ۸itﺣ۸ ou hﺣ۸tﺣ۸rogﺣ۸nﺣ۸itﺣ۸ de l’occupation de l’espace) et des facteurs de l’environnement. Nous comparerons ici uniquement les distributions parasites en fonction du sexe et de l’ﺣ۱ge de l’hﺣﺑte et en fonction des deux annﺣ۸es de prﺣ۸lﺣ۷vement (test U de Mann-Whitney).
2. Rﺣ۸sultats
2.1. Distibution de Ctenotaenia marmotae
La prﺣ۸valence observﺣ۸e pour cette espﺣ۷ce en fin de saison est maximale avec une abondance (ﺣ۸gale, de ce fait, ﺣ l’intensitﺣ۸ moyenne) trﺣ۷s ﺣ۸levﺣ۸e (tableau I). La prﺣ۸valence observﺣ۸e dans les Alpes italiennes au cours des mois de septembre et d’octobre (n = 7) est identique et l’abondance plus faible (33,6 + 51, ﺣ۸tendue 3-149) mais pas significativement diffﺣ۸rente de celle de notre ﺣ۸chantillon (Bassano et al. 1992). La comparaison avec d’autres donnﺣ۸es bibliographiques, sur la marmotte alpine ou d’autres espﺣ۷ces de marmottes, est rendue impossible par le fait que les auteurs ne distinguent pas dans leurs rﺣ۸sultats les valeurs obtenues ﺣ la fin de la saison d’activitﺣ۸ des hﺣﺑtes.
La distribution des frﺣ۸quences du parasite et les trois index d’agrﺣ۸gation (tableau II, figures 1 et 2) montrent une surdispersion nette de la population parasite dans la population-hﺣﺑte. L’index D prﺣ۸sente pourtant une valeur assez faible (0,49), et la divergence entre la droite et la courbe formﺣ۸e par les barres d’histogramme n’est pas trﺣ۷s forte (figure 2). Cependant du fait que la prﺣ۸valence est de 100 %, tous les hﺣﺑtes sont parasitﺣ۸sﺡ ; donc les parasites sont mieux rﺣ۸partis, quoique de faﺣ۶on trﺣ۷s irrﺣ۸guliﺣ۷re, dans la population-hﺣﺑte que lorsque la prﺣ۸valence est plus faible. De plus, les individus-hﺣﺑtes qui hﺣ۸bergent un trﺣ۷s grand nombre de parasites sont nombreuxﺡ (23 % portent plus de 200 vers). Ce qui explique valeur ﺣ۸levﺣ۸e de kﺡ : 1,083 .
Tableau Iﺡ : Index ﺣ۸pidﺣ۸miologiques parasitaires pour C. marmotae dans la population de marmottes de Bonneval-sur-Arc en fin d’ﺣ۸tﺣ۸ (n = 22)
|
Prﺣ۸valence + I.C. |
Abondance M + E.T. |
Intensitﺣ۸ moyenne M + E.T. |
Etendue |
|
100% + 4 |
96,4 + 96,3 |
96,4 + 96,3 |
1-309 |
I.C. = intervalle de confianceﺡ ; M = moyenneﺡ ; E.T. = ﺣ۸cart-type.
ﺡ
Tableau IIﺡ : Indices d’agrﺣ۸gation parasitaire pour C. marmotae dans la population en fin d’ﺣ۸tﺣ۸ (n = 22 ).
|
Rapport Variance/Moyenne (R) |
Paramﺣ۷tre binomial. nﺣ۸gatif (k3) |
Index de divergence (D) |
|
91,9 |
1,083 |
0,49 |
A cette pﺣ۸riode de l’annﺣ۸e, la majoritﺣ۸ des vers identifiﺣ۸s (97,9 %) prﺣ۸sentait la caractﺣ۸ristique des individus adultesﺡ : extrﺣ۸mitﺣ۸ caudale "carrﺣ۸e", traduisant la largeur des segments murs ﺣ۸mis.
Du fait de la grande taille deC. marmotae, la biomasse parasitaire qui en dﺣ۸coule est trﺣ۷s ﺣ۸levﺣ۸e (86 grammes pour l’individu 44 qui hﺣ۸bergeait 206 C. marmotae, soit 2 % du poids corporel de ce mﺣ۱le adulte de 4,4 kilogrammes, figure4). Les marmottes portent en moyenne une biomasse de 46,9 grammes de vers en fin de saison (soit un poids moyen pour un taenia de 0,4 gramme).

Figure 1ﺡ : Distribution de frﺣ۸quences de Ctenotaenia marmotae
en fin d’ﺣ۸tﺣ۸ sur le site de Bonneval-sur-Arc.

Figure 2ﺡ : Diagramme de Lorenz des pourcentages cumulﺣ۸s de Ctenotaenia marmotae
en fin d’ﺣ۸tﺣ۸ (la ligne droite indique la distribution uniforme idﺣ۸ale)
Le coefficient de corrﺣ۸lation r2 entre nombre de vers et biomasse parasitaire (n = 19) est de 0,31ﺡ ; il atteint 0,56 pour le sous-ﺣ۸chantillon ne comprenant que les valeurs infﺣ۸rieures ﺣ 200 vers/hﺣﺑte (n = 14) mais diminue ﺣ 0,12 pour l’autre partie (n = 5). La faiblesse de la corrﺣ۸lation au-dessus du seuil de 200 vers/hﺣﺑte, montre qu’une augmentation du nombre de vers prﺣ۸sents dans l’hﺣﺑte, si elle n’est pas liﺣ۸e ﺣ une augmentation correspondante de la biomasse, est due ﺣ une diminution de la taille des vers.
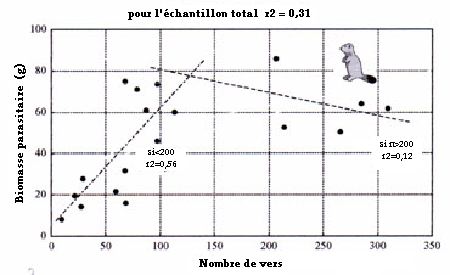
Figure 3ﺡ : Biomasse parasitaire de Ctenotaenia marmotae en fonction du nombre de vers.
(avec les droites de rﺣ۸gression linﺣ۸aire correspondant aux deux parties de l’ﺣ۸chantillon)
Ce phﺣ۸nomﺣ۷ne, "effet de surpeuplement" (crowding effect), est ﺣ۸tudiﺣ۸ depuis longtemps pour les cestodes (Read 1951, Euzeby 1966). Il est caractﺣ۸risﺣ۸ sur le plan morphologique par la diminution de la taille du nombre de vers, dont nous observons ici la consﺣ۸quence directe sur la biomasse parasitaire, et, sur le plan biologique, par un ralentissement de la croissance des cestodes ﺣ partir d’un certain seuil d’infestation. Le dﺣ۸lai d’acquisition de la maturitﺣ۸ sexuelle n’est pas affectﺣ۸ par ce phﺣ۸nomﺣ۷ne, car il est indﺣ۸pendant de la vitesse de croissance des vers, mais le nombre de segments ovigﺣ۷res ﺣ۸mis est considﺣ۸rablement rﺣ۸duit (Euzeby 1966).
Les valeurs de l’abondance et l’intensitﺣ۸ moyenne observﺣ۸es suivant le sexe et l’ﺣ۱ge de l’hﺣﺑte, et suivant les annﺣ۸es de prﺣ۸lﺣ۷vement ne montrent aucune diffﺣ۸rence significative (Mann-Whitney).
Distribution d’Ascaris laevis
La prﺣ۸valence d’A. laevis est ﺣ۸levﺣ۸e. L’abondance et l’intensitﺣ۸ moyenne sont plus faibles que pour Ctenotaenia marmotae (tableauIII). Les index calculﺣ۸s ﺣ partir des donnﺣ۸es de Bassano et al. (1992) sont significativement infﺣ۸rieursﺡ : prﺣ۸valence 28 %, abondance 3,6 + 7,2 et intensitﺣ۸ moyenne 12,5 + 9,2.
La distribution observﺣ۸e diffﺣ۷re d’une distribution normale (Kolmogorov-Smirnov, d = 0,24 p < 0,01).
Tableau IIIﺡ : Index ﺣ۸pidﺣ۸miologiques parasitaires pour Ascaris laevis dans la population de marmottes de Bonneval-sur-Arc en fin d’ﺣ۸tﺣ۸ (n = 22)
|
Prﺣ۸valence + I.C. |
Abondanceo M + E.T. |
Intensitﺣ۸ Moyenne M +E.T. |
Extrﺣ۹mes |
|
86% + 14 |
45,1 + 60,4 |
52,2 + 62,2 |
0-258 |
Tableau IVﺡ : Indices d’agrﺣ۸gation parasitaire pour Ascaris laevis dans la population de Bonneval-sur-Arc en fin d’ﺣ۸tﺣ۸ (n = 22 ).
|
Rapport Variance/Moyenne (R) |
Paramﺣ۷tre binomial. nﺣ۸gatif (k3) |
Index de divergence (D) |
|
77,3 |
0,786 |
0,56 |
Les trois index (tableau VI) et la figure 4 montrent une rﺣ۸elle agrﺣ۸gation de la population parasite dans la population-hﺣﺑte. L’index D est supﺣ۸rieur ﺣ celui de Ctenotaenia marmotae, et la divergence entre la droite et la courbe formﺣ۸e par les barres d’histogramme est plus grande (figure 5)ﺡ ; de mﺣ۹me le paramﺣ۷tre k indique une agrﺣ۸gation plus forte ﺣ cause de la prﺣ۸sence de deux individus-hﺣﺑtes porteurs d’un nombre extrﺣ۹me de vers. Au contraire, le rapport R est plus faible pour A. laevis que pour C. marmotae, car la majoritﺣ۸ des individus-hﺣﺑtes sont porteurs de moins de 80 vers.
Pour 6 individus-hﺣﺑtes le sexage des parasites a ﺣ۸tﺣ۸ rﺣ۸alisﺣ۸. La valeur du sex-ratio est de 0,37. Les femelles sont donc majoritaires en automne dans l’organisme-hﺣﺑte. De plus une grande partie d’entre elles est au stade adulte (prﺣ۸sence d’œufs, visibles par transparence dans le corps des individus).

Figure 4ﺡ : Distribution de frﺣ۸quence pour Ascaris laevis en fin d’ﺣ۸tﺣ۸ sur le site de Bonneval-sur-Arc.

ﺡ
Figure 5ﺡ : Diagramme de Lorenz des pourcentages cumulﺣ۸s d’Ascaris laevis
en fin d’ﺣ۸tﺣ۸ (la ligne droite indique la distribution uniforme idﺣ۸ale).

Figure 6: Biomasse parasitaire d’Ascaris laevis en fonction du nombre de vers
Ascaris laevis est, comme Ctenotaenia marmotae, un parasite de grande taille ﺣ l’ﺣ۸tat adulte. La biomasse parasitaire qu’il reprﺣ۸sente est parfois aussi impressionnanteﺡ : 80,3 grammes pour l’individu (32) de 4 kilogrammes hﺣ۸bergeant 66 parasites. Portant aussi 73,6 grammes de C. marmotae, la charge parasitaire totale de cet individu s’ﺣ۸levait donc ﺣ 3,9 % de son poids corporelﺡ ! Mais la figure 6 ne montre pas de relation linﺣ۸aire ﺣ۸troite entre le nombre de vers et la biomasse corporelle (r2 = 0,2). Au contraire, mis ﺣ part deux cas extrﺣ۹mes, toutes les biomasses parasitaires sont groupﺣ۸es autour des mﺣ۹mes valeurs ( < 20 grammes), quel que soit le nombre de vers.
La comparaison des valeurs de l’abondance et de l’intensitﺣ۸ moyenne observﺣ۸es suivant le sexe et l’ﺣ۱ge de l’hﺣﺑte, et suivant les annﺣ۸es de prﺣ۸lﺣ۷vement, test de Mann-Whitney, ne montre aucune diffﺣ۸rence significative pour toutes les combinaisons possibles. Au contraire, les mﺣ۱les et les femelles prﺣ۸sentent des valeurs trﺣ۷s proches d’abondance (p = 0,91) et d’intensitﺣ۸ moyenne (p = 0,83).
ﺡ
2.3. Distribution de Citellina alpina
La valeur de la prﺣ۸valence obtenue pour Citellina alpina est plus faible que celle des deux espﺣ۷ces prﺣ۸cﺣ۸dentesﺡ : elle atteint 67 % (tableau V). Le maximum de vers dﺣ۸nombrﺣ۸s a ﺣ۸tﺣ۸ de 230, alors que, dans la majoritﺣ۸ des cas, on observe la prﺣ۸sence de 5 ﺣ 25 parasites par hﺣﺑte sur le site ﺣ۸tudiﺣ۸ (figure 7).
Tableau Vﺡ : Index ﺣ۸pidﺣ۸miologiques parasitaires pour Citellina alpina dans la population de marmottes de Bonneval-sur-Arc en fin d’ﺣ۸tﺣ۸ (n = 22)
|
Prﺣ۸valence + I.C. |
Abondance M + E.T. |
Intensitﺣ۸ Moyenne M + E.T. |
Extrﺣ۹mes |
|
67% + 20 |
20,6 + 48,7 |
30,2 + 56,9 |
0-230 |
La distribution observﺣ۸e diffﺣ۷re d’une distribution Normale (Kolmogorov-Smirnov , d = 0,35 p < 0,01).
Les trois index d’agrﺣ۸gation (tableau VI) et la figure 7 illustrent la forte surdispersion de la population parasite dans la population-hﺣﺑte. L’index D prﺣ۸sente la valeur la plus ﺣ۸levﺣ۸e, et la divergence entre la droite et la courbe formﺣ۸e par les barres d’histogramme est encore plus concave que dans les deux cas prﺣ۸cﺣ۸dents (figure 8). De mﺣ۹me, le rapport R est plus ﺣ۸levﺣ۸ et le paramﺣ۷tre k plus faible que pour Ascaris laevis et Ctenotaenia marmotae. Ce degrﺣ۸ d’agrﺣ۸gation plus ﺣ۸levﺣ۸ est dﺣﭨ ﺣ la prﺣ۸sence, d’une part d’une majoritﺣ۸ d’individus porteurs de moins de 30 parasites et, d’autre part d’un seul individu prﺣ۸sentant un nombre de vers trﺣ۷s supﺣ۸rieur aux autres. En effet 50 % des vers sont portﺣ۸s par 95 % des hﺣﺑtes (figure 8).
Tableau VIﺡ : Indices d’agrﺣ۸gation parasitaire pour Citellina alpina dans la population de Bonneval-sur-Arc en fin d’ﺣ۸tﺣ۸ (n = 22 ).
|
Rapport Variance/Moyenne (R) |
Paramﺣ۷tre binomial. nﺣ۸gatif (k3) |
Index de divergence (D) |
|
109,9 |
0,492 |
0,73 |

Figure 7ﺡ : Distribution de frﺣ۸quence de Citellina alpina en fin d’ﺣ۸tﺣ۸.

Figure 8ﺡ : Pourcentages cumulﺣ۸s de Citellina alpina en fin d’ﺣ۸tﺣ۸
(la droite indique la distribution uniforme idﺣ۸ale).
Les vers adultes sont de petite taille (7 ﺣ 10 mm), donc la biomasse parasitaire qui dﺣ۸coule de leur prﺣ۸sence n’a pas ﺣ۸tﺣ۸ mesurﺣ۸e.
Dans tous les cas, le sexage des vers a montrﺣ۸ une forte prﺣ۸dominance des mﺣ۱lesﺡ : le sex-ratio est de 0,88. Dans la majoritﺣ۸ des infrapopulations, les parasites femelles prﺣ۸sents ﺣ۸taient des formes immaturesﺡ ; seul l’individu (53), qui prﺣ۸sentait la charge maximale en Citellina alpina ﺣ cette ﺣ۸poque de l’annﺣ۸e, arborait presque autant de femelles matures que de femelles immatures.
Comme prﺣ۸cﺣ۸demment, aucun des tests U ne signale de diffﺣ۸rence significative entre les diffﺣ۸rentes sﺣ۸ries, sexe et ﺣ۱ge de l’hﺣﺑte ainsi que l’annﺣ۸e de prﺣ۸lﺣ۷vement.
Discussionﺡ : Causes et consﺣ۸quences des distributions agrﺣ۸gﺣ۸es
Les distributions du nombre de parasites dans une population hﺣﺑte sont des "ﺡ entitﺣ۸s dynamiques plutﺣﺑt que statiques, gﺣ۸nﺣ۸rﺣ۸es par des forces opposﺣ۸es, certaines agissant pour crﺣ۸er la sur-dispersion et d’autres pour engendrer la sous-dispersionﺡ " (Anderson & May 1979ﺡ ; Anderson & Gordon 1982ﺡ ; figure 9).
| Facteurs qui gﺣ۸nﺣ۷rent la sous-dispersion | Facteurs qui gﺣ۸nﺣ۷rent la sur-dispersion |
| 1. Mortalitﺣ۸ parasite | Hﺣ۸tﺣ۸rogﺣ۸nﺣ۸itﺣ۸ dans la sensibilitﺣ۸ hﺣﺑte ﺣ l’infestation |
| 1. Processus densitﺣ۸-dﺣ۸pendants | 2. Structures spatiales et sociales de la population hﺣﺑte |
| 3. Mortalitﺣ۸ hﺣﺑte induite par le parasite | 3. Hﺣ۸tﺣ۸rogﺣ۸nﺣ۸itﺣ۸ dans la capacitﺣ۸ hﺣﺑte ﺣ tuer les parasites par rﺣ۸ponse immunologique ou autre |
| 4. Mode de transmission du parasite | |
| 5. Reproduction directe du parasite dans l’hﺣﺑte |
Figure 9ﺡ : Facteurs ﺣ l’origine de la dispersion des parasites (Anderson & Gordon 1982, modifiﺣ۸).
Cet aspect dynamique nous a fait choisir de dresser un "ﺣ۸tat des lieux" de la distribution des parasites de la marmotte alpine sur un site dﺣ۸limitﺣ۸ et dans un intervalle de temps rﺣ۸duit. Nous avons ainsi pu montrer un ﺣ۸tat de sur-dispersion ou d’agrﺣ۸gation diffﺣ۸rent pour les trois espﺣ۷ces d’helminthes, Ctenotaenia marmotae, Ascaris laevis et Citellina alpina, dans un ﺣ۸chantillon d’individus-hﺣﺑtes.
3.1. Origines de l’agrﺣ۸gation
Les processus ﺣ l’origine des patrons de dispersion des parasites sont liﺣ۸s ﺣ l’hﺣ۸tﺣ۸rogﺣ۸nﺣ۸itﺣ۸ de la relation de la population-hﺣﺑte avec la population-parasite (Combes 1995). Les "causes" de l’agrﺣ۸gation peuvent ﺣ۹tre rangﺣ۸es dans deux catﺣ۸goriesﺡ : celles qui dﺣ۸pendent du parasite et celles qui dﺣ۸pendent de l’hﺣﺑte. Dans chacune de ces catﺣ۸gories, se distinguent ﺣ nouveau deux types de mﺣ۸canismesﺡ : ceux liﺣ۸s ﺣ la rencontre hﺣﺑte/parasite et ceux liﺣ۸s au dﺣ۸veloppement et ﺣ la survie du parasite dans l’hﺣﺑte.
3.1.1. Facteurs liﺣ۸s aux parasites
Le mode de transmission du parasite est un ﺣ۸lﺣ۸ment dﺣ۸terminant de la rencontre avec ses hﺣﺑtes potentiels. On distingue les parasites "ﺡ contagieuxﺡ ", ﺣ transmission directe (contact physique des hﺣﺑtes entre eux ou avec leurs dﺣ۸jections) ou indirecte par le milieu (sans dﺣ۸placement par un vecteur) et les parasites transmissibles (transmission par vecteurﺡ ; Cote & Poulin 1995, Fromont 1997).
Dans le groupe des parasites "ﺡ contagieuxﺡ ", certaines espﺣ۷ces, comme Ascaris laevis et Citellina alpina, prﺣ۸sentent un cycle direct (pas de stade dispersant). Le parasite ne peut donc pas aller ﺣ la rencontre de son hﺣﺑte. Pour augmenter ses "chances" de transmission, il doit augmenter le nombre de ses propagules ﺣ۸mises dans les fﺣ۷ces de l’hﺣﺑte, puis dissﺣ۸minﺣ۸es dans le milieu extﺣ۸rieur ou sur le pelage de l’hﺣﺑte. Le parasite doit aussi augmenter leur capacitﺣ۸ de rﺣ۸sistance dans le milieu extﺣ۸rieur. Ce mode de transmission favorise donc les parasites ﺣ longue durﺣ۸e de vie, ﺣ fﺣ۸conditﺣ۸ ﺣ۸levﺣ۸e et dont les propagules sont bien rﺣ۸sistantes dans l’environnement. Ces mﺣ۸canismes augmentent la densitﺣ۸ des stades infestants de parasites dans le milieu extﺣ۸rieur aprﺣ۷s maturation. Les pertes au cours de la transmission sont ﺣ۸normes. Chez le lagopﺣ۷de, un stade libre sur un million du parasite Trichotrongylus tenuis devient adulte (Dobson & Hudson 1992)ﺡ ! D’autre part, le taux de rencontre entre les hﺣﺑtes ﺣ۸ventuels et les stades infestants est fortement influencﺣ۸ par la distribution spatiale de ces derniers (Keymer & Anderson 1979)ﺡ : la distribution, elle-mﺣ۹me agrﺣ۸gﺣ۸e, des stades infestants dans le milieu extﺣ۸rieur peut permettre l’acquisition des parasites en "lots" plus ou moins grands (Combes 1995).
La biologie d’Ascaris laevis, avec une longﺣ۸vitﺣ۸ probable de plusieurs mois et une bonne survie des œufs dans le milieu extﺣ۸rieur, en particulier dans la neige, explique donc en partie la sur-dispersion observﺣ۸e. De plus, la distribution des propagules dans le milieu est agrﺣ۸gﺣ۸e, car les fﺣ۷ces, qui les renferment, sont regroupﺣ۸es dans les nombreuses latrines externes dissﺣ۸minﺣ۸es sur chaque territoire.
Pour Citellina alpina, la rﺣ۸sistance des œufs est probablement de courte durﺣ۸e. Mais le manque de connaissances sur sa longﺣ۸vitﺣ۸ et sa fﺣ۸conditﺣ۸ ne permettent pas d’ﺣ۸valuer l’efficacitﺣ۸ de sa transmission et d’expliquer les mﺣ۸canismes de rencontre ainsi, que l’origine de l’agrﺣ۸gation observﺣ۸e.
ﺡ
Dans le groupe des espﺣ۷ces transmissibles, les parasites, tels que Ctenotaenia marmotae, prﺣ۸sentent des cycles de vie indirects avec deux hﺣﺑtes (ou plus), en gﺣ۸nﺣ۸ral mobiles, pour complﺣ۸ter leur dﺣ۸veloppement. La transmission se fait par des liens de type "prﺣ۸dateur-proie"ﺡ avec une phase d’infestation de l’hﺣﺑte-intermﺣ۸diaire (ingestion d’œufs prﺣ۸sents dans les fﺣ۷ces par les Oribates coprophages dans le cas de C. marmotae), suivie de la contamination de l’hﺣﺑte-dﺣ۸finitif (ingestion des Oribates infestants par les marmottes). Les contraintes imposﺣ۸es par ces liens "prﺣ۸dateur-proie" ne sont probablement pas dﺣ۸savantageuses pour la stabilitﺣ۸ et la persistance des populations-parasites puisqu’elles sont frﺣ۸quentes dans les cycles de vie des helminthes (Keymer & Anderson 1979).
Enfin, l’infestation multiple des hﺣﺑtes-intermﺣ۸diaires conduit ﺣ l’infestation des hﺣﺑtes-dﺣ۸finitifs par "lots"ﺡ : l’agrﺣ۸gation du parasite, dans le premier lot, participe ﺣ la crﺣ۸ation de l’agrﺣ۸gation dans le second. Par contre, alors qu’un parasite semble rare dans son hﺣﺑte intermﺣ۸diaire, une telle raretﺣ۸ ne s’observe pas nﺣ۸cessairement chez l’hﺣﺑte dﺣ۸finitif (Hair 1975). Ainsi, chez Ctenotaenia marmotae,, sa prﺣ۸valence chez l’hﺣﺑte intermﺣ۸diaire est estimﺣ۸e ﺣ 0,4 % alors qu’elle est de 100 % chez l’hﺣﺑte dﺣ۸finitif en automne (Ebermann 1976). La combinaison de ces processus chez C. marmotae, explique l’infestation des individus examinﺣ۸s. L’efficacitﺣ۸ des stratﺣ۸gies de transmission du parasite est ﺣ l’origine d’une dispersion moins forte des vers de cette espﺣ۷ce que l’agrﺣ۸gation montrﺣ۸e pour les deux autres helminthes.
Le phﺣ۸nomﺣ۷ne de favorisationﺡ : processus ﺣ۸thologique adaptatif modifiant les positions spatio-temporelles du parasite et de sa cible dans un sens qui augmente la probabilitﺣ۸ de leur rencontre a ﺣ۸tﺣ۸ dﺣ۸fini par Combes (1991). Ainsi, les cystacanthes de l’acanthocﺣ۸phale Polymorphus paraxodus provoquent l’expression de comportements de l’amphipode hﺣﺑte-intermﺣ۸diaire qui augmentent significativement ses chances d’ﺣ۹tre ingﺣ۸rﺣ۸s par le canard hﺣﺑte-dﺣ۸finitif (Holmes & Bethel 1972). Un tel phﺣ۸nomﺣ۷ne serait ﺣ rechercher chez les hﺣﺑtes intermﺣ۸diaires de Ctenotaenia marmotae pour expliquer l’ﺣ۸cart de prﺣ۸valence observﺣ۸ entre les deux hﺣﺑtes, intermﺣ۸diaire et dﺣ۸finitif. En fait, l’influence de la manipulation comportementale de l’hﺣﺑte par le parasite est probablement trﺣ۷s sous-estimﺣ۸e (Thompson & Kavaliers 1994ﺡ , Combes 1995).
Une fois que le parasite a atteint son hﺣﺑte, il doit ﺣ۹tre capable de vivre et de se dﺣ۸velopper dans cet organisme jusqu’ﺣ sa reproduction sexuﺣ۸e dans l’hﺣﺑte dﺣ۸finitif, sa maturation ou sa multiplication asexuﺣ۸e dans l’hﺣﺑte intermﺣ۸diaire. Pour cela la sﺣ۸lection s’exerce au niveau des parasites, pour favoriser ceux qui possﺣ۷dent la meilleure capacitﺣ۸ ﺣ s’adapter ﺣ l’environnement hﺣﺑte, notamment ﺣ ﺣ۸chapper ﺣ l’immunitﺣ۸ de celui-ci (Combes 1995). Cette adaptation nﺣ۸cessaire conduit le parasite ﺣ rﺣ۸duire le nombre d’hﺣﺑtes dans lesquels il peut vivreﺡ : la restriction de la dimension hﺣﺑte de la niche ﺣ۸cologique d’un parasite est la rﺣ۷gle (Rhodes 1994, Combes 1995), mﺣ۹me si certaines espﺣ۷ces parasites sont capables de se dﺣ۸velopper, avec plus ou moins de succﺣ۷s, dans un grand nombre d’espﺣ۷ces hﺣﺑtes. Il peut ainsi arriver qu’un hﺣﺑte soit rﺣ۸ceptif ﺣ peu d’espﺣ۷ces de parasites, mais les hﺣ۸berge en abondance. Ainsi, chez deux espﺣ۷ces sympatriques de crabes, hﺣﺑtes intermﺣ۸diaires d’helminthes parasitesﺡ : l’une possﺣ۷de une faune parasitaire riche mais trﺣ۷s peu abondante, alors que l’autre prﺣ۸sente une communautﺣ۸ parasitaire pauvre mais trﺣ۷s abondante (Bush et al. 1993).
Parmi les trois helminthes de la communautﺣ۸ parasitaire de la marmotte alpine, recensﺣ۸s en abondance sur le site de Bonneval-sur-Arc, Ctenotaenia marmotae et Ascaris laevis sont stﺣ۸noxﺣ۷nes et Citellina alpina oﺣﺁoxﺣ۷ne. Leur abondance dans les autres populations ou espﺣ۷ces-hﺣﺑtes est rarement connue, ﺣ l’exception des ﺣ۸tudes chez la marmotte alpine oﺣﺗ elles forment aussi la structure principale des communautﺣ۸s parasitaires observﺣ۸es (Sabatier 1989, Bassano et al., 1992, Manfredi et al., 1992, Preleuthner et al., 1996). La spﺣ۸cialisation des trois helminthes prﺣ۸sents dans les infracommunautﺣ۸s de marmottes alpines ﺣ۸tudiﺣ۸es favorise l’agrﺣ۸gation dans le sens oﺣﺗ les formes infestantes ingﺣ۸rﺣ۸es, en quantitﺣ۸s variables, par la marmotte ont de bonnes chances de se dﺣ۸velopper dans leur hﺣﺑte dﺣ۸finitif.
3.1.2. Facteurs liﺣ۸s ﺣ l’hﺣﺑte
Certains individus-hﺣﺑtes peuvent offrir une meilleure "qualitﺣ۸ d’habitat" que d’autres (Combes 1995), favorisant l’agrﺣ۸gation des individus-parasites. Ainsi, un habitat de bonne qualitﺣ۸ pour un parasite est un hﺣﺑte que le parasite peut rencontrer facilement et avec lequel la compatibilitﺣ۸ est possible.
La nature des contacts entre individus-hﺣﺑtes sains et infestﺣ۸s est dﺣ۸terminante pour la transmission du parasite ﺣ l’hﺣﺑte, en particulier pour les parasites contagieux. Les structures spatiale et sociale des populations-hﺣﺑtes sont donc susceptibles de favoriser ou non l’agrﺣ۸gation parasitaire.
Les structures spatiales sont les plus ﺣ۸tudiﺣ۸es. Fromont (1997) distingue les populations homogﺣ۷nes, dans lesquelles tous les individus ont la mﺣ۹me probabilitﺣ۸ de rencontre, et les populations hﺣ۸tﺣ۸rogﺣ۷nes, dans le cas contraire. Les colonies de marmottes prﺣ۸sentent des structures spatiales hautement hﺣ۸tﺣ۸rogﺣ۷nesﺡ : l’organisation en mﺣ۸tapopulations, ensemble de colonies interconnectﺣ۸es, correspond ﺣ un cas extrﺣ۹me d’hﺣ۸tﺣ۸rogﺣ۸nﺣ۸itﺣ۸. Au sein d’une colonie donnﺣ۸e, la rﺣ۸partition des marmottes en groupes familiaux, territoriaux est susceptible de favoriser la transmission des parasites, par rapport ﺣ une distribution rﺣ۸guliﺣ۷re (Blower & Roughgarden 1989ﺡ , Grosholz 1993). L’importance des charges parasitaires dﺣ۸nombrﺣ۸es montre que l’utilisation des mﺣ۹mes latrines par les membres du groupe, le toilettage mutuel, la cohabitation dans les mﺣ۹me terriers sont des sources de transmission parasitaire trﺣ۷s efficaces. De plus, la taille des groupes, parfois importante chez la marmotte alpine (10 individus ou plus), peut accroﺣ؟tre encore le risque d’infestation. En effet, Cote & Poulin (1995) ont montrﺣ۸ l’existence d’une corrﺣ۸lation positive entre la prﺣ۸valence du parasite et la taille du groupe hﺣﺑte, dans une mﺣ۸ta-analyse regroupant six ﺣ۸tudes d’espﺣ۷ces sociales. Ils n’ont au contraire pas mis en ﺣ۸vidence de relation entre charge parasitaire et taille du groupe, mﺣ۹me chez les espﺣ۷ces hﺣﺑtes sﺣ۸dentaires pour lesquelles on pouvait attendre une corrﺣ۸lation positive. L’absence de connaissances sur la composition des groupes d’oﺣﺗ sont issues les marmottes de Bonneval-sur-Arc ne nous permet pas d’ﺣ۸tudier cet aspect.
Chez la marmotte la structure spatiale se mﺣ۹le ﺣ l’organisation sociale : les relations entre les individus dﺣ۸pendent de leur statut hiﺣ۸rarchique. Les interactions sociales, amicales ou agonistiques, entre individus, le choix du partenaire sexuel pour les individus dominants, favorisent les contacts. Ces relations peuvent donc influencer ou ﺣ۹tre influencﺣ۸es par le parasitisme.
L’ﺣ۸tude de la relation entre les niveaux d’infestation par des ectoparasites et la socialitﺣ۸, en conditions naturelles, chez la marmotte alpine (Arnold & Lichtenstein 1993) comme chez la marmotte ﺣ ventre jaune (Van Vuren 1996) ne montre pas d’augmentation significative de la charge en ectoparasites avec la taille des groupes sociaux. Le nombre moyen d’Echinonyssus blanchardi, diffﺣ۸rent d’un groupe ﺣ l’autre, n’est pas corrﺣ۸lﺣ۸ avec la densitﺣ۸ de marmottes alpines sur un territoire donnﺣ۸ (Arnold & Lichtenstein 1993). Les charges en ectoparasites des marmottes ﺣ ventre jaune ne sont pas plus ﺣ۸levﺣ۸es chez les individus coloniaux que chez ceux qui ne le sont pas, ni plus ﺣ۸levﺣ۸es chez les mﺣ۱les polygynes (censﺣ۸s avoir plus de contacts avec les autres membres du groupe, en particulier avec les femelles) que chez les monogames (Van Vuren 1996). Ne connaissant pas la structure sociale des groupes d’origine de notre ﺣ۸chantillon, nous ne pouvons apporter aucune information nouvelle concernant les relations entre le statut social des individus et la charge parasitaire pour chacun des helminthes recensﺣ۸s. De plus, il est trﺣ۷s difficile de prﺣ۸dire dans quel sens peuvent se faire ces relations, puisque les deux hypothﺣ۷ses proposﺣ۸es par Combes (1995) "ﺡ plus ﺣ۸levﺣ۸ le statut, moins abondant le parasiteﺡ " et "ﺡ plus ﺣ۸levﺣ۸ le statut, plus abondant le parasiteﺡ " sont vﺣ۸rifiﺣ۸es tour ﺣ tour suivant les systﺣ۷mes hﺣﺑte-parasite ﺣ۸tudiﺣ۸s. Par exemple, une ﺣ۸tude expﺣ۸rimentale montre que l’infestation des souris par le nﺣ۸matode Heligmosomoides polygyrus peut empﺣ۹cher les mﺣ۱les d’accﺣ۸der ﺣ la dominance comportementale ﺣ partir d’une certaine charge parasitaire (Freeland 1981).
En ce qui concerne la structure dﺣ۸mograph ique, Arnold & Lichtenstein (1993) montrent qu’il n’y a pas de diffﺣ۸rence significative de la charge parasitaire entre mﺣ۱le et femelle. L’analyse statistique des donnﺣ۸es de notre ﺣ۸chantillon ne dﺣ۸tecte pas de diffﺣ۸rence entre les sexes, ni pour la prﺣ۸valence ni pour l’abondance et l’intensitﺣ۸ moyenne.
Ce sont ici les dimensions immunitaires et personnelles des individus-hﺣﺑtes qui sont susceptibles de favoriser ou non l’agrﺣ۸gation. L’immunitﺣ۸, sous l’influence des caractﺣ۷res gﺣ۸nﺣ۸tiques, conduit ﺣ rendre les individus rﺣ۸sistant aux parasites de faﺣ۶on trﺣ۷s inﺣ۸gale. Le phﺣ۸notype de chaque individu, et donc sa capacitﺣ۸ ﺣ rﺣ۸sister ou non aux parasites, est sous l’influence de l’histoire de vie, prﺣ۸sente ou passﺣ۸e, de chaque animal (Anderson & May 1978). Ainsi il a ﺣ۸tﺣ۸ montrﺣ۸ que les communautﺣ۸s de parasites se modifient qualitativement et quantitativement au cours de la vie des individus-hﺣﺑtes, en particulier chez une espﺣ۷ce de poisson mﺣ۸diterranﺣ۸en (Saad-Fares & Combes 1992). Cet exemple est certes ﺣ۸loignﺣ۸ de celui de la marmotte et notre ﺣ۸chantillon n’a pas permis de mettre en ﺣ۸vidence une relation entre l’ﺣ۱ge des hﺣﺑtes et la richesse spﺣ۸cifique ou la charge parasitaire. Indﺣ۸pendamment de l’ﺣ۱ge, tout ﺣ۸vﺣ۸nement, susceptible d’affaiblir l’hﺣﺑte, peut favoriser l’infestation.
Chez la marmotte alpine, un ﺣ۸vﺣ۸nement de cette nature intervient au moins une fois dans la vie des hﺣﺑtesﺡ : la dispersion. Les individus subadultes sont soit chassﺣ۸s de leur territoire d’origine, soit chassent eux-mﺣ۹mes le dominant du mﺣ۹me sexe de leur groupe d’origine pour prendre sa place, soit partent d’eux-mﺣ۹mes chercher une place de dominant dans un autre groupe. Ces diffﺣ۸rentes stratﺣ۸gies sont ﺣ۸tudiﺣ۸es au niveau comportemental (Magnolon et al. 1997), mais le parasitisme peut aussi participer ﺣ leur dﺣ۸terminisme. Holekamp (1986), chez une espﺣ۷ce d’ﺣ۸cureuil, et Van Vuren (1996), chez la marmotte ﺣ ventre jaune, ont ﺣ۸mis l’hypothﺣ۷se que les individus se dispersent si le risque parasitaire est ﺣ۸levﺣ۸, se coupant ainsi du contact social. Ce dernier a en effet montrﺣ۸ que la charge en ectoparasites a tendance ﺣ ﺣ۹tre plus ﺣ۸levﺣ۸e chez les marmottes qui sont en pﺣ۸riode de dispersion que chez les autres, bien que l’auteur prﺣ۸cise que la diffﺣ۸rence n’est pas statistiquement significative.
Le stress et la nutrition peuvent aussi participer ﺣ la crﺣ۸ation d’une dispersion plus ou moins forte des parasites entre les hﺣﺑtes.
Notre ﺣ۸chantillon est ﺣ۸videmment de taille insuffisante pour aborder l’influence de ces divers facteurs. La complexitﺣ۸ des structures de populations de marmottes laisse penser que l’influence respective des diffﺣ۸rents facteurs sur l’agrﺣ۸gation observﺣ۸e ne pourrait ﺣ۹tre mise en ﺣ۸vidence que grﺣ۱ce ﺣ une ﺣ۸tude pluridisciplinaire, en ﺣ۸thologie et parasitologie. Or la rﺣ۸alisation des bilans parasitaires indispensables nﺣ۸cessite de sacrifier des individus dans les groupes sociaux ﺣ۸tudiﺣ۸s, ce qui perturbe l’organisation sociale de ceux-ci et rend les observations ﺣ۸thologiques inopﺣ۸rantes.
3.2, Consﺣ۸quences de l’agrﺣ۸gation
Anderson (1978) a ﺣ۸tabli queﺡ : les patrons agrﺣ۸gﺣ۸s ou sur-dispersﺣ۸s du nombre de parasites par hﺣﺑtes, largement observﺣ۸s dans les habitats naturels, agissent en augmentant la rﺣ۸gulation densitﺣ۸-dﺣ۸pendante ﺣ la fois des populations hﺣﺑtes et parasites. Anderson & Gordon (1982) prﺣ۸cisent que "ﺡ c’est dans les quelques hﺣﺑtes porteurs de la majoritﺣ۸ des parasites que les processus densitﺣ۸-dﺣ۸pendants exercent leur influence rﺣ۸gulatrice, soit via la suppression de la fﺣ۸conditﺣ۸ ou de la survie parasitaire, soit via l’influence du parasite sur la survie et la fﺣ۸conditﺣ۸ de l’hﺣﺑteﺡ ".
La rﺣ۸gulation est un mﺣ۸canisme de dynamique de population, qui s’applique partout, dﺣ۷s l’instant oﺣﺗ le milieu dans lequel vit la population abrite des ressources inﺣ۸vitablement limitﺣ۸es (Barbault 1997). L’hﺣﺑte du parasite est nﺣ۸cessairement un milieu limitﺣ۸ et seul un processus de rﺣ۸gulation s’oppose ﺣ l’accumulation a priori illimitﺣ۸e d’individus-parasites par recrutement dans le milieu extﺣ۸rieur.
Combes (1995) dﺣ۸finit la rﺣ۸gulation commeﺡ : "ﺡ le maintien d’une population ﺣ un effectif infﺣ۸rieur ﺣ sa capacitﺣ۸ de croissance thﺣ۸orique, par des mﺣ۸canismes densitﺣ۸-dﺣ۸pendantsﺡ ". Elle se manifeste dans une population ﺣ partir d’un certain seuil, ﺣ۸videmment variable suivant le parasite et l’hﺣﺑte en cause. Les modﺣ۷les rﺣ۸alisﺣ۸s pour dﺣ۸crire les relations entre populations-parasites et populations-hﺣﺑtes (Crofton 1971a et 1971b, Anderson & May 1978, May & Anderson 1978) montrent que la rﺣ۸gulation de la population—hﺣﺑte par un parasite nﺣ۸cessite, soit l’agrﺣ۸gation de la distribution des parasites, soit une augmentation de l’effet pathogﺣ۷ne plus rapide que l’augmentation de la charge parasitaire.
3.2.1. Rﺣ۸gulation des populations—hﺣﺑtes par les parasites :
A l’ﺣ۸chelle des individus, le parasite est, par dﺣ۸finition, considﺣ۸rﺣ۸ comme dﺣ۸favorable, responsable de la maladie, voire de la mort de son hﺣﺑte. Par contre, ﺣ l’ﺣ۸chelle d’une population ou d’une espﺣ۷ce, le mﺣ۹me parasite peut en permettre la survie. Par exemple l’effet pathogﺣ۷ne du parasite (au sens large, incluant virus et bactﺣ۸ries) participe ﺣ la rﺣ۸gulation d’une population trop dense, oﺣﺗ la compﺣ۸tition, spatiale ou alimentaire, devient trop forte, en crﺣ۸ant une dﺣ۸croissance brusque du nombre d’individus (Holmes 1982). Mais le parasite peut agir de faﺣ۶on moins visibleﺡ : ﺣ۸tant en compﺣ۸tition avec son hﺣﺑte pour ses ressources, en particulier alimentaires dans le cas d’un parasite du tractus digestif, il peut en affecter la fﺣ۸conditﺣ۸ et la survie, mﺣ۹me si aucune pathologie n’est visible. L’influence du parasite peut se noyer dans le bruit de fond de tous les autres facteurs qui affectent la fﺣ۸conditﺣ۸ et la survie de l’hﺣﺑte (Combes 1995) et est souvent difficile ﺣ mettre en ﺣ۸vidence, surtout pour les macroparasites. Pourtant, en conditions semi-naturelles, la survie des individus de l’espﺣ۷ce Apodemus sylvaticus parasitﺣ۸s par le nﺣ۸matode Heligmosomoides polygyrus est infﺣ۸rieure ﺣ celle des individus sains (Gregory 1991).
Il ressort des ﺣ۸tudes, concernant les helminthes en particulier, que les effets rﺣ۸gulateurs des populations-parasites sur les populations-hﺣﺑtes deviennent visibles quand les hﺣﺑtes sont soumis ﺣ des parasites auxquels ils ne sont pas prﺣ۸parﺣ۸s et/ou quand la compﺣ۸tition alimentaire devient telle que les individus-hﺣﺑtes sont affaiblis par la malnutrition (Gulland 1992).
Chez la marmotte alpine, nous avons recherchﺣ۸ un effet rﺣ۸gulateur de l’infestation parasitaire sur le site de la Grande Sassiﺣ۷re, en particulier sur l’accroissement pondﺣ۸ral des individus au cours de la saison, sans succﺣ۷s du fait de l’insuffisance de captures en automne. La difficultﺣ۸ ﺣ mettre en ﺣ۸vidence une telle relation tient au fait que l’augmentation de poids des marmottes est un processus plurifactoriel, et que, sur le site d’ﺣ۸tude, les sources de nourriture des marmottes sont suffisamment abondantes et de bonne qualitﺣ۸ pour ﺣ۸viter la compﺣ۸tition alimentaire et que les marmottes sont bien "habituﺣ۸es" ﺣ leurs parasites.
ﺡ
3.2.2. Rﺣ۸gulation des populations-parasites par les hﺣﺑtes :
Une infrapopulation parasite ne s’accroﺣ؟t pas indﺣ۸finiment dans son individu-hﺣﺑte. Un processus rﺣ۸gulateur intervient donc sur l’infrapopulation, ﺣ la fois de la part des autres parasites prﺣ۸sents et de la part de l’hﺣﺑte. La rﺣ۸gulation par l’interaction des parasites entre eux fait intervenir les processus de compﺣ۸tition, intra- et interspﺣ۸cifique que nous envisagerons dans la discussion suivante.
Nous avons vu que le niveau moyen d’infestation parasitaire (prﺣ۸valence et abondance) de la marmotte est trﺣ۷s ﺣ۸levﺣ۸ et variable suivant les espﺣ۷ces parasites. Les mﺣ۸canismes dﺣ۸veloppﺣ۸s par l’hﺣﺑte dﺣ۸finitif pour limiter l’infestation parasitaire sont d’origine immunitaire (Grundmann et al. 1976ﺡ ; Anderson& May 1978) et la sﺣ۸vﺣ۸ritﺣ۸ de la rﺣ۸ponse immunologique par l’hﺣﺑte est liﺣ۸e au niveau de stimulation antigﺣ۸nique reﺣ۶ue, c’est-ﺣ -dire au nombre de parasites prﺣ۸sents dans l’infracommunautﺣ۸. Anderson & Michel (1977) ont ﺣ۸tabli pour Ostertagia ostertagi que, une telle rﺣ۸ponse, d’origine humorale ou cellulaire, tend ﺣ augmenter le taux de mortalitﺣ۸ dans la population parasite et/ou ﺣ rﺣ۸duire son potentiel reproductif. Chez la souris ﺣ pattes blanches (Peromyscus maniculatuss), un seul gﺣ۷ne autosomal contrﺣﺑle le dﺣ۸veloppement de l’immunitﺣ۸ contre le cestode Hymenolepis citelli. (Wassom et al. (1974)
L’ﺣ۸tude des processus immunitaires mis en œuvre par les marmottes pour contrﺣﺑler leur infestation parasitaire demande des investigations que nous n’avons pas rﺣ۸alisﺣ۸es dans cette ﺣ۸tude. La nature et l’efficacitﺣ۸ des mﺣ۸canismes en cause sont probablement trﺣ۷s diffﺣ۸rentes suivant l’espﺣ۷ce parasite, comme en tﺣ۸moignent les fortes variations d’abondancesﺡ : l’immunitﺣ۸ mise en œuvre contre Ctenotaenia marmotae serait-elle moins efficace que celle destinﺣ۸e ﺣ Citellina alpina, l’abondance du second ﺣ۸tant, en fin de saison, nettement moins forte que celle du premierﺡ ?
ﺡ
4. Conclusion
Les infracommunautﺣ۸s d’helminthes parasites de la marmotte alpine sont structurﺣ۸es et prﺣ۸dictibles dans le sens oﺣﺗ elles semblent ﺣ۹tre le rﺣ۸sultat de processus complexes qui ne doivent que peu de chose au hasard et oﺣﺗ leur organisation n’est pas diffﺣ۸rente d’une annﺣ۸e ﺣ l’autre.
Le comportement de la marmotte, son organisation sociale et spatiale jouent des rﺣﺑles trﺣ۷s importants dans la crﺣ۸ation des patrons agrﺣ۸gﺣ۸s de distribution des infrapopulations.
Le mode de transmission des parasites, contagieux ou transmissibles, favorise et entretient cette agrﺣ۸gation.
Les consﺣ۸quences de la distribution agrﺣ۸gﺣ۸e des parasites sont difficiles ﺣ mettre en ﺣ۸vidence car, en l’absence de pathologie visible, les effets du parasitisme sur la survie et la fﺣ۸conditﺣ۸ de l’hﺣﺑte sont difficiles ﺣ distinguer des autres facteurs qui les affectent.
La marmotte semble donc ﺣ۹tre un "habitat de bonne qualitﺣ۸" pour ses helminthes parasites, qui, eux, sont capables de "tirer partie" de ses particularitﺣ۸s comportementales.
ﺡ
ﺡ
ﺡ
Bibliographie
Anderson R.M. & May R.M. 1978. Regulation and stability of host-parasite population interactions. I. regulatory processes. Journal of Animal Ecology, 47: 219-247.
Anderson R.M. & May R.M. 1979. Population biology of infectious diseasesﺡ : Part I. Nature, 280ﺡ 361-367.
Anderson R.M. & Gordon D.M. 1982. Processes influencing the distribution of parasite numbers within host populations with special emphasis on parasite-induced host mortalities. Parasitology, 85: 373-398.
Arnold W. & Lichtenstein A.V. 1993. Ectoparasite loads decrease the fitness of alpine marmot (Marmota marmota) but are not a cost of sociality. Behavioral Ecology, 41:36-39.
Bassano B., Sabatuer B., Rossi L. & Macchi E. 1992. Parasitic fauna of the digestive tract of Marmota marmota in the western alps. In First International Symposium on Alpine Marmot (Marmota marmota) and genus Marmota, Bassano B., Durio P., Gallo Orsi U. & Macchi E., 13-24.
Blower S.M. & Roughgarden J. 1989. Parasites detects host spatial pattern and densityﺡ : a field experimental analysis. Oecologia, 78: 138-141.
Bush A.O., Heard R.W.J. & Overstreet R.M. 1993. Intermediate hosts as source communities. Canad Journal of Zoology, 71: 1358-1363.
Combes C. 1995. Interactions durablesﺡ : Ecologie et ﺣ۸volution du parasitisme. Masson, Paris, 524 p.
Cote I.M. & Poulin R. 1995. Parasitism and group size in social animals: a meta-analysis. Behav. Ecol., 6: 159-165.
Crofton H.D. 1971a. A quantitative approach to parasitism. Parasitoloy 62: 179-193.
Crofton H.D. 1971b. A model of host-parasite relationships. Parasitoloy, 63: 343-364.
Dobson A.P. 1985. The population dynamics of competition between parasites. Parasitology., 91: 317-347.
Dobson A.P. & Hudson P.J. 1992. Regulation and stability of free-living host-parasite systemﺡ : Trichostrongylus tenuis in red grose. II Population models. Journal of Animal Ecology, 61: 487-498.
Ebermann E. 1976. Oribatids as intermediate hosts of Ctenotaenia marmotae (Frﺣﭘlich, 1802), a tapeworm parasitic in marmots. Zeitschrift fur Parasitenkunde, 50ﺡ (3): 303-312.
Euzeby J. 1966. Les maladies vermineuses des animaux domestiques et leurs incidences sur la pathologie humaine. In Maladies dues aux Plathelminthes, tome II, Vigot frﺣ۷res ed., Paris, 663 p.
Fromont E. 1997. Analyse comparative de la transmission de cinq virus dans des populations de chats domestiques (Felis catus L.). Thﺣ۷se Doc. Universitﺣ۸, Lyon, 139 p.
Frontier S. & Pichon-Viale 1998. Ecosystﺣ۷mes, structures, fonctionnement, ﺣ۸volution. Masson ed., Paris, 447 p.
Grosholz E.D. 1993. The influence of habitat heterogeneity on host-pathogen population dynamics. Oecologia, 96: 347-353.
Hair J.D. 1975. The structure of the intestinal helminths communities of lesser scaup (Aythya affinis). Alberta Univ.
Holekamp K.E. 1986. Proximal causes of natal dispersal in Belding’s ground squirrel (Spermophilus beldingi). Ecol. Monogr., 56: 365-391.
Holmes J.C. & Bethel W.M. 1972. Modification of intermediate host behavior by parasites. In Behavioral aspects of parasite transmission, vol. 51 (suppl.1), J. of the Zoological Linnean Society, Canning & Wright eds., 123-149.
Keymer A.E. & Anderson R.M. 1992. The dynamics of infection of Tribolium confusum by Hymenolepis diminuaﺡ : the influence of infective-stage density and spatial distribution. Parasitology, 79: 195-207.
Lorenz M.O. 1905. Methods for measuring the concentration of wealth. American Statistical Association, 9: 209-219.
Magnolon Sﺡ ;, Allainﺣ۸ D. & Coulon J. 1997. A study of the proximal factors affecting natal dispersal in the alpine marmot (Marmota marmota). In Holarctic marmots as a factor of biodiversity, Rumiantsev V.Yu., Nikol’ski A.A., Brandler O.V., Moscow, 166.
Manfredi M.T., Zanin E. & Rizzoli A.P. 1992. Helminth community on Alpine marmots. In First International symposium on Alpine marmot (Marmota marmota) and genus Marmota, Bassano B.ﺡ et al. edsﺡ ., 203-207.
May R.M. & Anderson R.M. 1978. Regulation and stability of host-parasite interactions. II. Destabilizing processes. Journal of Animal Ecology, 47: 249-267.
Pennycuick L. 1971. Frequency distributions of parasites in a popualtion of three-spined stickelbacks, Gasterosteus aculeatus L., with particular reference to the negative binomial distribution. Parasitology, 63: 389-406.
Poulin R. 1993. The disparity between observed and uniform distributionﺡ : a new look at parasite aggregation. Intern. J. Parasitol., 26(7): 937-944.
Poulin R. 1995. Misconceptions about the measurement of aggregationﺡ : a reply to Ploeger. Intern. J. Parasitol.,, 25(7): 863-864.
Preleuthner M., Prosl H., Bregmann A., & Pinsker W. 1996. Infestation by endoparasites in different Austrian populations of Marmota marmota. In Biodiversity in Marmots, Le Berre M., Ramousse R. & Le Guelte L. eds., 111-112.
Read C.P. 1951. The "ﺡ crowding effectﺡ " in tapeworm infections. The Journal of Parasitology, 37: 174-178.
Rhode K. 1994. Niche restriction in parasitesﺡ : proximate and ultimate causes. Parasitology, 109: S69-S84.
Saad-Fares A. & Combes C. 1992. Abundance/host size relationship in a fish trematode comminity. Journal of Helminthology, 66: 187-192.
Sabatier B. 1989. Les parasites de la marmotte alpineﺡ : ﺣ۸tude dans les Alpes franﺣ۶aises et synthﺣ۷se bibliographique. Thﺣ۷se de Doctorat Vﺣ۸tﺣ۸rinaire, 178 p.
Scherrer B. 1984. Biostatisque. Gaﺣ۹tan Morin, Quebec, 850 p.
Shaw D.J. & Dobson A.P. 1995 Patterns of macroparasite abundance and aggregation in wildlife populationsﺡ : a quantitative review. Parasitology, 111: S111-S113..
Scott M.E. 1997. Temporal changes in aggregationﺡ : a laboraory study. Parasitology, 94: 583-595.
Thompson S.N. & Kavaliers M. 1994. Phsyiological bases for parasite-induced alterations of host behaviour. Parasitology, 109: S119-S138.
Van Vuren D. 1996. Ectoparasites, fitness and social behavior of yellow-bellied marmots. Ethology, 102: 686-694.