![]()
Marmota vancouverensis, Swarth, occupaient surtout les prairies
subalpines (1000 et 1400 mètres d'altitude) de l'île de Vancouver.
Ces dernières années, elles ont colonisé les
clairières de déforestation (<700 mètres d'altitude),
et ont disparu de certains habitats naturels. La population actuelle est
à la fois faible (100-200 individus) et très concentrée.
Ses taux de reproduction sont peu élevés : portées de
petite taille (![]() = 3.36, SD = 0.83, n = 36), maturité sexuelle tardive
(1ère reproduction = 4,00 ans, ET = 0,82, n = 13) et longs
intervalles entre portées sans reproduction (= 1.83 ans, ET = 0.76,
n = 6). Prédation et échec d'hibernation sont les
principales causes de mortalité. Les marmottes de clairières ont
moins de marmottons et présentent des taux de survie plus faibles que
les marmottes des habitats naturels. Les clairières, "puits" à
population, consomment plus de marmottes qu'elles n'en produisent. La
sylviculture peut, ainsi, ralentir ou prévenir la reconquête de
certains habitats historiques.
= 3.36, SD = 0.83, n = 36), maturité sexuelle tardive
(1ère reproduction = 4,00 ans, ET = 0,82, n = 13) et longs
intervalles entre portées sans reproduction (= 1.83 ans, ET = 0.76,
n = 6). Prédation et échec d'hibernation sont les
principales causes de mortalité. Les marmottes de clairières ont
moins de marmottons et présentent des taux de survie plus faibles que
les marmottes des habitats naturels. Les clairières, "puits" à
population, consomment plus de marmottes qu'elles n'en produisent. La
sylviculture peut, ainsi, ralentir ou prévenir la reconquête de
certains habitats historiques.
Mots-clés : Marmotte de l'île de Vancouver, Marmota vancouverensis,
démographie, sylviculture, métapopulation, protection.
Vancouver Island Marmots (Marmota vancouverensis: Swarth) normally
inhabit sub-alpine meadows at 1000-1400 metres elevation. In recent years
marmots colonized habitats created by clearcut logging of forests above 700 m
in elevation, but disappeared from some natural habitats. The current
population is small (100-200 individuals) and highly concentrated. This marmot
exhibits low reproductive rates, with small litters (
![]() = 3.36, SD = 0.83, n = 36), late sexual maturity ( age at first
reproduction = 4.00, SD = 0.82, n = 13) and a lengthy non-reproductive
interval between litters (= 1.83 years, SD = 0.76, n = 6). Predation and
unsuccessful hibernation are the most important causes of mortality. Marmots
inhabiting recently logged habitats produce fewer young and show significantly
lower survival rates. Clearcut habitats may act as population "sink" by
consuming more marmots than they produce. Forestry may ultimately slow or
prevent recolonization of some historic habitats.
= 3.36, SD = 0.83, n = 36), late sexual maturity ( age at first
reproduction = 4.00, SD = 0.82, n = 13) and a lengthy non-reproductive
interval between litters (= 1.83 years, SD = 0.76, n = 6). Predation and
unsuccessful hibernation are the most important causes of mortality. Marmots
inhabiting recently logged habitats produce fewer young and show significantly
lower survival rates. Clearcut habitats may act as population "sink" by
consuming more marmots than they produce. Forestry may ultimately slow or
prevent recolonization of some historic habitats.
Key-words: Vancouver Island Marmot, Marmota vancouverensis,
demography, forestry, metapopulations, conservation.

Dès 1980, les marmottes ont colonisé les habi-tats déboisés entre 800-1100 mètres (33% de la population en 1984 et >50% en 1994). Cependant, cela n'a entraîné ni son augmentation numérique ni celle de son aire de distribution, qui, en fait, a diminué ces dernières décades. Les marmottes occupent maintenant moins de 50 sites sur 15 montagnes. Toutes les colonies, sauf une, sont situées entre 5 lignes de partage des eaux voisines dans le centre-sud de l'île de Vancouver (Figure 1).
La relation entre Marmota vancouverensis et la sylviculture est méconnue. D'une part, cette dernière a créé de grandes surfaces d'habitats potentiels dont cer-tains ont été colonisés par les marmottes. D'autre part, les marmottes ont disparu de nombreux sites historiques naturels malgré leur maintien en état. Ce travail porte sur le statut global de la population et compare l'évolu-tion démographique des marmottes dans les prairies subalpines naturelles et les clairières.
Les sites Haley Lake (1100 mètres d'altitude) et Green Mountain (1400 m) sont des prairies pentues (30 à 45deg.), exposées au sud ou sud-ouest, libres d'arbres du fait des glissements de neige et avalanches. Ils sont occupés depuis longtemps (relevés de 1915 à Haley Lake et de 1954 à Green Mountain). Le site Vaughan Road, situé à 1 kilomètre à l'ouest de la colonie Haley Lake, a été déboisé entre 1974 et 1978 (940 m). Il est escarpé, avec de nombreux talus, rochers et zones érodées ; libre de végétation au début de l'étude, il est occupé depuis par A. sitchensis et des conifères de régénération. Les marmottes y ont été observées pour la première fois en 1985. Le site Pat Lake (920 m) est une cuvette escarpée, exposée au nord, entourant un lac peu profond de 16 km au sud de la colonie Haley Lake et à 2 km au nord-est de Mount Whymper. Déboisé entre 1978 et 1979, les marmottes y ont été découvertes en 1985 et sont présentes dans plusieurs parcelles d'habi-tats naturels. La végétation est comparable à celle de Vaughan Road, avec des communautés dominantes de Lupinus et Anaphalis. Le site SL, à 0,9 km et 260 m en dessous d'un site naturel ou existe une petite colonie, exposé au sud (980 m) sur le flanc de Mount Landalt, a été déboisé en 1977, et les premières marmottes y ont été observées en 1991.
Les tailles des populations des sites intensi-vement suivies ont été basées sur plus de 5 dénombre-ments annuels, incluant une forte proportion d'animaux marqués. L'effort d'échantillonnage de ces colonies a été enregistré en journées d'étude (visite d'une personne ou plus à une colonie par jour), en jours par personne (une personne sur un site durant une session matinale ou vespérale, par beau temps) et en jours-pièges (un piège utilisé sur un site durant un jour, compte non tenu du nombre de déplacements du piège parmi les sites).
La survie suivant l'âge et le sexe a été estimée par les méthodes des tables standard de survie (survie standardisée, Caughley 1977). Mes données, incluant la dispersion, sous-estiment les taux réels de survie. Les échantillons de marmottons marqués étant réduits, j'ai estimé leurs taux de survie en comparant leur nombre et celui des un-ans observés l'année suivante. Les animaux marqués revus ont été utilisés pour estimer les taux de survie des autres classe d'âge (Bryant sous presse).
Les populations des colonies intensivement étu-diées fluctuent ; la plupart montre un déclin depuis le début de l'étude (Table 1). L'essentiel de la variation de la taille globale de la population est le résultat des variations des nombres de juvéniles et de un-ans survi-vants. En moyenne, 62% des marmottes observées dans les colonies étudiées de 1987 à 1995 ont été finalement marquées à l'oreille, soit plus de 13% de la population probable de M. vancouverensis de ces années.
Les femelles peuvent se reproduire dès 3 ans (âge moyen de 1ère reproduction = 4 ans ; ET = 0,23 ; écart = 3-5). Cinq femelles ont engendré des portées multiples (une, 3 portées ; et 2 autres, des portées des années consécutives), représentant 59% (n = 37) de tous les marmottons des sites naturels. Aucune femelle de clairière n'a produit plus d'une portée. La fécondité à un âge donné a été relativement stable avec l'accrois-sement de l'âge, et similaire quelle que soit le type d'habitat des animaux de même âge (Fig. 3).
Malgré l'impossibilité d'identifier les pères, le comportement et le faible nombre d'animaux permet-tent d'émettre quelques conclusions sur la structure so-ciale. Malgré un biais en faveur des femelles du nombre d'adultes, dans la plupart des colonies, la plupart des années, le nombre de mâles est suffisant pour suspecter l'existence d'unités monogames. Cependant, à 3 occa-sions, des portées multiples ont été engendrées dans des colonies ne comprenant qu'un seul mâle adulte, suggé-rant l'existence de la polygamie chez cette espèce. Ces mâles ont engendré 2 portées à Green Mountain(1988) et à Haley Lake (1994), ou 3 portées dans la nouvelle colonie Sherk Lake.
Les taux de survie, après groupement des classes d'âge, sont significativement inférieurs dans les clai-rières (50 sur 104 ; 48%) à ceux des prairies (84 sur 129; 65%; [[chi]]2 = 6.84, 1 ddl, p <0.01). Bien que la faiblesse des échantillons en fonction de l'âge gêne l'analyse, la tendance générale a été une réduction de la survie de 15-20% de toutes les classes d'âge et de sexe (Fig. 4). Des femelles d'habitats naturels sont représentées dans les quatre classes d'âge supérieures alors qu'il n'y a aucune femelle de clairière (Fig. 3). Le fait essentiel est que les taux de reproduction nette des colonies (Ro) sont très inférieurs dans les clairières (Ro = 0.25) à ceux des habitats naturels (Ro =0.72).
Huit groupes familiaux comprenant des marmottons ont disparu entre fin août et début mai. Dans tous les cas, les terriers principaux sont soit inoccupés au printemps, soit occupés exclusivement par de nou-veaux adultes, non marqués (7 cas dans les clairières et 1 cas dans les prairies naturelles). Dans d'autres cas, les adultes ont survécu mais les marmottons ont disparu. La survie de ces derniers a été très faible en 1989-90 (13 sur 31 individus), en 1990-91 (0 sur 8), en 1994-95 (8 sur 27) et élevée en 1991-92 (10 sur 16) et en 1993-94 (17 sur 23). Mes données n'excluent pas la possibilité que certains marmottons disparus se soient dispersés, bien que cela soit peu probable, compte-tenu de l'absence de données concernant de jeunes dispersants.
Une seule mort imputable à l'étude s'est pro-duite (1992) suite à l'hyperthermie d'un mâle adulte lors de son transport précédant l'implantation d'un radioémetteur. Un mâle d'un-an a été tué dans un piège armé par un visiteur. Enfin, 3 marmottes ont été abattues par des vandales (Janz et al. 1994).
Les marmottes de l'île de Vancouver sont rares, surtout du fait de leurs habitats parcellisés et dis-persés. Le profond changement de climat et le développement de la forêt qui ont suivi la dernière période gla-ciaire n'ont laissé que de petites parcelles de prairies appropriées aux marmottes, typiquement dans des cu-vettes avalancheuses ou sur les versants voisins (Milko 1984). M. vancouverensis peut fournir un bon exemple de dynamique naturelle d'une métapopulation consti-tuée de petites colonies au milieu de forêts ou d'habitats alpins non appropriés (Bryant 1990). Certaines caracté-ristiques comportementales de M. vancouverensis, son niveau élevé de socialité (Heard 1977) et sa capacité à réaliser de longs déplacements (Bryant & Janz sous presse), sont le résultat de son histoire évolutive.
La déforestation au-dessous de 800 m des années 1960 à 1980 a créé des milliers d'hectares d'habitats potentiels, offrant aux marmottes des sols propices à l'activité fouisseuse, des plantes variées à consommer, et une abondance de points de surveillance. Ceci, combiné aux conditions climatiques régionales favori-sant des taux de survie élevés et une bonne reproduc-tion, a permis aux marmottes d'occuper de nouveaux sites et d'accroître leur nombre au début des années 1980 (Bryant & Janz sous presse). Cependant, ce processus n'a entraîné ni une expansion de l'aire de distribution ni, malgré de nouveaux sites potentiels, un fort accroissement numérique.
La principale différence entre les marmottes des habitats naturels ou des clairières est une réduction de la survie de ces dernières. Au cours de l'étude (9 ans), aucune femelle des 3 colonies de clairière n'a atteint l'âge de 5 ans. Ces femelles ne peuvent atteindre la même performance reproductive que celles des habitats naturels. Je suggère que ces colonies de clairières se maintiennent grâce à une immigration continue de nouvelles femelles, et que les clairières fonctionnent comme des "puits" démographiques (Pulliam 1988).
M. vancouverensis est actuellement confinée sur moins de 15 montagnes, surtout au centre-sud de l'île de Vancouver (Bryant & Janz sous presse). Une interpréta-tion de la distribution actuelle est basée sur l'idée que les habitats historiques de l'espèce ne sont plus appro-priés du fait de l'évolution climatique et des modifica-tions de la couverture végétale qui y sont associées (Nagorsen et al. sous presse). Cette hypothèse découle logiquement de celle de Milko (1984), qui suggérait que l'invasion des prairies subalpines par les arbres limite les habitats de marmottes ou entraîne une réduction de la nourriture disponible (Martell & Milko 1986). Dans ce cadre, les habitats anciennement occupés par les mar-mottes ont changé d'une façon qualitative, et l'espèce est naturellement confinée à une région géographique, aux conditions climatiques et végétales satisfaisantes, en déclin. La découverte récente de 5 sites préhistoriques de fossiles de marmottes, tous largement hors de la zone centrale de la distribution actuelle, soutien cette hypo-thèse (Nagorsen & Keddie 1994, Nagorsen et al. sous presse). L'hypothèse d'une régulation climatique s'ac-corde aux vues de Thomas (1994), qui soutien que de nombreuses espèces rares dépendantes des conditions du milieu, disparaissant localement là où les conditions ne sont plus convenables, et colonisent des sites où les conditions s'améliorent.
La seconde interprétation est basée sur les notions de métapopulation et de source d'effondrement (Bryant 1990). Dans ce cadre, les habitats historiques de marmottes sont vacants car les extinctions locales n'ont pas été équilibrées par des recolonisations du fait de l'altération des éléments essentiels de la structure méta-populationnelle (dispersion réussie ou "connectivité") par l'activité humaine. Les preuves d'une telle structure chez M. vancouverensis comprennent l'observations de fluctuations des populations locales, d'extinctions et de recolonisations (Bryant 1990, Bryant & Janz sous presse). La preuve d'une connectivité réduite est ambi-guë ; l'idée que des habitats récents mais de mauvaise qualité, voisins des prairies naturelles, entravent la dispersion, est en accord avec les données génétiques (Bryant 1990), et la concentration spatiale des événe-ments de colonisation (Bryant & Janz sous presse), mais des preuves d'une dispersion à longue distance dans un paysage modifié existent aussi (Bryant sous presse, Bryant & Janz sous presse).
Le choix entre ces hypothèses ne sera pas aisé. Le problème fondamental est celui de l'absence de témoin expérimental (il n'y a pas de ligne de partage des eaux, déboisée, où les marmottes soient présentes et où des niveaux "normaux" de connectivité du paysage puissent être mesurés). L'hypothèse climatique, aussi, n'est pas vérifiable directement. Bien que des méthodes permettent de mesurer la stabilité des habitats (dendro-chronologie, par exemple), il est impossible d'attribuer l'absence de marmottes de zones particulières à une relation de cause à effet basée sur la variation du climat.
La question de savoir pourquoi les marmottes n'occupent plus certaines zones est fondamentale pour leur protection. Si les modification climatiques ont altéré les habitats historiques, les efforts de restauration des colonies échoueront et il y a fort peu de choses que les gestionnaires puissent faire pour restaurer les popu-lations. Au contraire, si l'altération de la connectivité du paysage est due à l'activité humaine, alors les déplace-ments peuvent être un moyen simple et économique de rétablir les populations de marmottes (par exemple, Ramousse et al. 1992), et les perspectives d'un éventuel "rétablissement" de l'espèce sont alors excellentes.
Je suis redevable à L. DYCK, D. MILNE & J. ROBIN, qui m'ont assisté sur le terrain, et à l'équipe de réhabilitation des marmottes de l'île de Vancouver, qui m'a assuré de son aide inlassable pour la recherche et la gestion de M. vancouvernsis. Les financements ont été assurés par B.C. Habitat Conservation Fund, B.C. Ministry of Environment, Lands and Parks, World Wildlife Fund (Canada), Cowi-chan Valley Field Naturalists Society et Forest Alliance de B.C. et Canadian wildlife service. Mac Millan Bloedel Limited and Timber West m'ont autorisé l'accès aux propriétés privées. La permission de capture des marmottes dans la Réserve Ecologique de Haley Lake m'a été donnée par Ecological Reserves Unit of B.C.
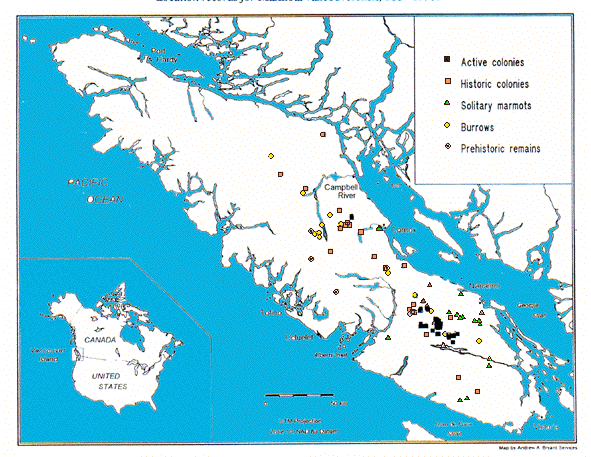
Les colonies actives sont celles confirmées par l'équipe inventoriale depuis 1980. Les colonies historiques sont antérieures à cette date. La population actuelle est concentrée dans les parties supérieures des fleuves Namaido, Cowichan, Chemainus, Cameron et Nitinat, et plus de 75% des animaux ont été découverts sur une surface de 20 km2. Active colonies include those confirmed by inventory crews since 1980. Historic colonies were reported prior to this date. The current population is concentrated within the headwaters of the Nanaimo, Cowichan, Chemainus, Cameron and Nitinat river systems, with >75% of known animals found within an area of approximately 20 km2. From Bryant & Janz (in press).
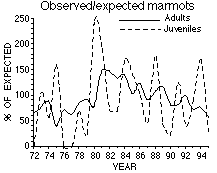 | 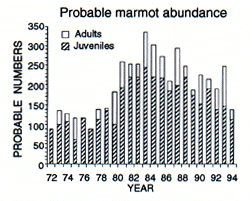 |
Les rapports observés/attendus (A) sont basés sur la moyenne d'abondance de chaque site, seuls les sites observés chaque année sont pris en compte. Le nombre probable de marmottes (B) a été estimé après correction basée sur l'effort de comptage en excluant les habitats de clairière les années précédant la colonisation. La population actuelle (1995) est inférieure à 200 animaux.
Observed/expected ratios (A) were based on average abundance for each site, using only those sites counted in any year. Probable marmot numbers (B) were estimated by applying a correction factor based on count effort, and by excluding clearcut habitats for years prior to colonization. The current (1995) population contains fewer than 200 animals.
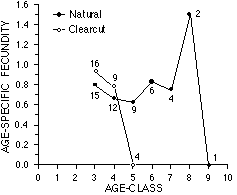
Les données sont calculées à partir des comptages des jeunes nés de marmottes marquées, assumant une sex-ratio des marmottons équilibrée. La taille des échantillons est donnée. la fécondité est généralement stable avec l'âge. Les femelles des clairières présentent un succès reproducteur au cours de leur vie fortement réduit (2,27 marmottons/femelle, comparé à 4,4 marmottons/femelle dans les habitats naturels). D'après Bryant (sous presse).
Data were calculated from counts of infants born to ear-tagged marmots assuming 1:1 juvenile sex ratio. Sample sizes are shown. Fecundity is generally stable with increasing age. Females in clearcut habitats show considerably reduced lifetime reproductive success (2.27 juveniles/female, compared with 4.4 juveniles/female in natural habitats). From Bryant (in press).
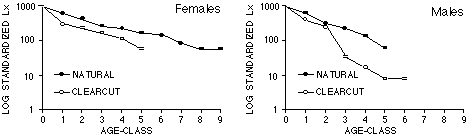
Les taux de survie des jeunes ont été estimés à partir des comptages de jeunes non marqués et des un-an survivants. Les autres taux ont été estimés à partir des animaux marqués. La tendance globale est une faible réduction (15-20%) de la survie des animaux occupant les clairières.
Infant survival rates were estimated from counts of untagged infants and surviving yearlings. Other age-specific rates were estimated from tagged animals. The overall trend is a subtle reduction (15-20%) in survival of animals living in logged habitats.
| 1987 | 1988 | 1989 | 1990 | 1991 | 1992 | 1993 | 1994 | 1995 | SD | |||
| Natural colonies | ||||||||||||
| Green Mountain | ||||||||||||
| N of adults | 5 | 6 | 3 | 5 | 5 | 2 | 1 | 4 | 3 | 3.78 | 1.64 | |
| N of yearlings | 0 | 0 | 4 | 0 | 0 | 1 | 2 | 2 | 0 | 1.00 | 1.41 | |
| N of juveniles | 3 | 6 | 0 | 0 | 4 | 3 | 3 | 0 | 0 | 2.11 | 2.20 | |
| Haley Lake | ||||||||||||
| N of adults | 12 | 14 | 12 | 12 | 8 | 7 | 9 | 5 | 7 | 9.56 | 3.05 | |
| N of yearlings | 4 | 1 | 5 | 0 | 3 | 6 | 0 | 10 | 3 | 3.56 | 3.21 | |
| N of juveniles | 2 | 12 | 0 | 3 | 9 | 0 | 12 | 10 | 0 | 5.33 | 5.32 | |
| Cumulative statistics | ||||||||||||
| Sampling (person-days) | 54 | 43 | 19 | 13 | 12 | 22 | 28 | 28 | 40 | 28.78 | 14.30 | |
| Sampling (trap-days) | 110 | 76 | 66 | 40 | 37 | 39 | 32 | 41 | 6 | 51.89 | 27.09 | |
| Tagging rate | 0.42 | 0.51 | 0.71 | 0.75 | 0.59 | 0.58 | 0.63 | 0.74 | 0.92 | 0.67 | 0.16 | |
| Accumulated tagged animals | 11 | 22 | 26 | 30 | 34 | 39 | 47 | 58 | 59 | |||
| Clearcut colonies | ||||||||||||
| Vaughan Road | ||||||||||||
| N of adults | 6 | 8 | 2 | 7 | 4 | 2 | 6 | 4 | 2 | 4.56 | 2.30 | |
| N of yearlings | 3 | 0 | 3 | 0 | 0 | 0 | 0 | 2 | 0 | 0.89 | 1.36 | |
| N of juveniles | 0 | 6 | 4 | 0 | 0 | 0 | 5 | 3 | 0 | 2.00 | 2.50 | |
| Pat Lake | ||||||||||||
| N of adults | 10 | 11 | 10 | 9 | 6 | 7 | 6 | 5 | 8.00 | 2.27 | ||
| N of yearlings | 5 | 3 | 0 | 0 | 2 | 3 | 3 | 2 | 2.25 | 1.67 | ||
| N of juveniles | 7 | 4 | 0 | 3 | 5 | 3 | 3 | 0 | 3.13 | 2.36 | ||
| F19 | ||||||||||||
| N of adults | 3 | 4 | 4 | 4 | 3.75 | 0.50 | ||||||
| N of yearlings | 0 | 0 | 0 | 3 | 0.75 | 1.50 | ||||||
| N of juveniles | 0 | 0 | 11 | 0 | 2.75 | 5.50 | ||||||
| Cumulative statistics | ||||||||||||
| Sampling (person-days) | 18 | 36 | 11 | 14 | 8 | 55 | 81 | 53 | 76 | 39.11 | 28.30 | |
| Sampling (trap-days) | 38 | 90 | 22 | 48 | 10 | 72 | 126 | 84 | 49 | 59.89 | 36.52 | |
| Tagging rate | 0.11 | 0.36 | 0.48 | 0.53 | 0.38 | 0.67 | 0.61 | 0.89 | 1.00 | 0.55 | 0.29 | |
| Accumulated tagged animals | 1 | 14 | 22 | 25 | 27 | 38 | 46 | 65 | 68 | |||
| Natural colonies | Clearcut colonies | |||||||
| Variable | +/- SD | range | n | +/- SD | range | n | Test statistic1 | p value |
| Litter size 1 | 3.35 +/- 0.93 | 2-5 | 20 | 3.38 +/- 0.72 | 2-5 | 16 | U (20,16) = 173 | 0.636 |
| Ratio of male:female juveniles 2 | 13:20 | 33 | 15:21 | 36 | c2 = 0.037 | >0.75 | ||
| Annual fecundity 3 | 1.27 +/- 1.77 | 0-5 | 49 | 1.28 +/- 1.73 | 0-5 | 29 | U (49,29) = 702 | 0.919 |
| Age of first reproduction 4 | 4.33 +/- 0.33 | 3-5 | 6 | 3.71 +/- 0.76 | 3-5 | 7 | U (6,7) = 30 | 0.173 |
| N of years between litters | 1.83 +/- 0.76 | 1-3 | 6 | - | 0 | - | - | |
1 Données incluant les 8 portées des femelles non marquées ; Data include 8 litters produced by untagged females. 2 Données incluant les 25 animaux capturés comme un an.; Data include 25 animals originally captured as yearlings. 3 Incluant les marmottons des deux sexes des femelles reproductrices marquées ; Includes infants of both sexes produced by ear-tagged females of reproductive age. 4 Données provenant des femelles capturées comme marmotton, un an ou deux ans ; Data from females originally captured as infants, yearlings or 2 year-olds.
The Vancouver Island marmot (Marmota vancouverensis: Swarth 1911) is endemic to Vancouver Island, British Columbia (Nagorsen 1987). It is the rarest member of the genus Marmota, with a current (1995) population estimated at 100-200 individuals (Bryant & Janz in press). M. vancouverensis was listed as "endangered" by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) in 1979, and by the Province of British Columbia in 1980 (Munro et al. 1985). A Recovery Team was established in 1988, and implementation of a recovery program began in 1993 (Janz et al. 1994).
Beginning in 1981, Vancouver Island marmots colonized new habitats created by logging of forests between 800 and 1100 metres elevation. By 1984, 33% of the known population was found in clearcuts, and by 1994 this figure had risen to >50%. Curiously, this did not result in expanded geographic range or greatly in-creased numbers. Indeed, the geographic range of the species has shrunk in the past few decades, and mar-mots now occupy fewer than 50 sites on 15 mountains, with all but 1 colony situated within 5 adjacent water-sheds on south-central Vancouver Island (Figure 1).
The relationship between M. vancouverensis and forestry is therefore somewhat mysterious. Forestry has created large areas of potential habitat, some of which have been colonized by marmots. On the other hand, marmots have disappeared from many historic natural habitats, despite their remaining in pristine condition. In this paper I report on overall population status, and compare demographic trends of marmots occupying natural sub-alpine meadow environments and logging clearcuts.
The Haley Lake and Green Mountain sites are steep (30deg. to 45deg.) south or southwest-facing meadows kept free of trees by snow-creep and avalanches at 1100 and 1420 metres elevation respectively. Both sites have a long history of marmot occupancy, with records dating from 1915 (Haley) and 1954 (Green). The Vaughan Road site is located 1 kilometre west of the Haley Lake colony, in an area that was logged between 1974 and 1978 (940 metres ASL). Marmots were first observed there in 1985. The topography is steep, with numerous talus slopes, boulders and eroded areas that were essentially bare of vegetation when the study began, but which are now dominated by A. sitchensis and regenerating conifers. The Pat Lake site is a steep north-facing bowl surrounding a shallow lake 16 kilometres southeast of the Haley Lake colony and 2 kilometres northeast of Mount Whymper, where marmots occur in several patches of natural habitat. Elevation at Pat Lake is 920 metres. The site was clearcut between 1978 and 1979. Marmots were first discovered there in 1985. As with Vaughan Road, vegetation is a complex of regenerating conifers, Alnus thickets, shrubs, grasses and Lupinus and Anaphalis-dominated communities. The Sherk Lake site is a south facing slope at 980 metres elevation on the southern flank of Mount Landalt. The area was logged in 1977, and marmots were first reported there in 1991. The site is 0.9 km distant from, and 260 metres lower than, a site where a small colony exists in natural habitat.
Population sizes given for intensively-studied sites were based on >5 annual counts, and included a large proportion of ear-tagged animals. Sampling effort at intensively-studied colonies was recorded in terms of field-days (a visit to a colony by 1 or more persons on 1 day), person-days (1 person on site during a morning or evening session in good weather) and trap-days (1 trap used at one site during 1 day, regardless of how many times the trap was moved among placements).
Age and sex-specific survival was estimated using standard life-table methods and presented as standardized survivorship (Caughley 1977). My data include dispersal and thus underestimate actual survi-val rates. Because sample sizes of tagged infants were small, I estimated infant survival rates by comparing counts of infants observed with counts of yearlings observed 1 year later. Re-sightings of marked animals were used to estimate survival rates for other age-classes (Bryant in press).
Populations at intensively-studied colonies fluc-tuated, although most show a decline since the study began (Table 1). Most of the variation in total population size was produced by changing numbers of juveniles and surviving yearlings. On average, 62% of all mar-mots observed at the study colonies from 1987 to 1995 were eventually ear-tagged, representing >13% of the probable population of M. vancouverensis during these years.
Females can reproduce as 3 year-olds, but mean age of first reproduction was 4.00 (SD = 0.23, range = 3-5). Five females produced multiple litters, including 1 animal which produced 3 litters during the study, and 2 females which produced litters in consecutive years. Cumulatively, these 5 females produced 59% (n = 37) of all infants observed at the natural sites. No female produced more than 1 litter in a clearcut site. Age-specific fecundity was relatively stable with increasing age, and values for natural and clearcut habitats were similar for animals of equal age (Figure 3).
It was impossible to identify male parents, but behavior and small numbers of animals permit some reasonable inferences about social structure. Although numbers of adults were apparently biased towards females, in most years, and for most colonies, numbers of males were sufficient for me to suspect single-male:single-female breeding groups. However, on three occasions, multiple litters were produced at colonies which contained only a single adult male, suggesting that polygamy does occur in this species. Single males produced 2 litters in 1988 at Green Mountain, 2 litters in 1994 at Haley Lake, and 3 litters in 1994 at the newly colonized Sherk Lake site.
When age-sex classes were pooled, survival rates were significantly lower in clearcuts (50 of 104; 48%) than in natural meadows (84 of 129; 65%; [[chi]]2 = 6.84 with 1 df, p <0.01). Although small age-specific samples hinder analysis, the overall trend was a 15-20% reduction in survival across all age-sex classes (Figure 4). Females in natural habitats were confirmed alive in 4 oldest age-classes that contained no clearcut females (Figure 3). The essential arithmetic is that colony net reproductive rates (Ro) were much lower in clearcut habitats (Ro = 0.25) than natural habitats (Ro =0.72).
Unsuccessful hibernation was also important. At least 8 family groups including juveniles disappeared between late August and early May observations. In all cases, habitual "home" burrows showed no evidence of use by marmots in the spring, or were occupied by new, untagged adults exclusively (7 cases in clearcuts and 1 case in a natural meadow). In other cases, adults survived but juveniles disappeared. Juvenile survival was particularly low in 1989-90 (13 of 31 animals), 1990-91, (0 of 8), and 1994-95 (8 of 27). High juvenile survival was observed in 1991-92 (10 of 16) and 1993-94 (17 of 23). My data do not exclude the possibility that some disappearing juveniles dispersed, although I consider this to be unlikely based on the absence of data showing "young" immigrants.
A single research mortality occurred in 1992, after an adult male overheated while being moved prior to implantation of a radio transmitter. A yearling male was killed in 1994 after a trap was set and left open by unknown visitors to the site. At least 3 marmots have been shot by vandals (Janz et al. 1994).
Vancouver Island marmots are rare primarily because their natural habitats are small and scattered. Pronounced climatic change and forest succession following the most recent glacial period left only small patches of grass-forb meadow habitat suitable for mar-mots, typically in avalanche bowls or adjacent to talus slopes (Milko 1984). M. vancouverensis may provide a good example of natural "metapopulation dynamics", in which the entire population consists of small colonies located in a matrix of unsuitable forest or alpine habitat (Bryant 1990). I believe that some behavioral attributes of M. vancouverensis, including its high level of social-ity (Heard 1977) and ability to make long-distance dispersal movements (Bryant & Janz in press), are the inevitable result of this evolutionary history.
Logging of forests above 800 metres during the 1960s, 1970s and 1980s created thousands of hectares of potential habitat, much of which offered suitable soil conditions for burrowing, a variety of grasses and forbs to eat, and a abundance of logs, stumps and boulders to serve as lookout spots. This, possibly combined with regional weather conditions that promoted good reproduction and survival rates, allowed marmots to colonize new sites and expand in numbers during the early 1980s (Bryant and Janz in press). However, this did not result in expanded geographic range or, given the amount of newly-created potential habitat, in greatly increased numbers.
The major difference among marmots living in natural and clearcut habitats was reduced survival. In 9 years of field-work at 3 clearcut colonies, no female was observed to produce more than a single litter, and only a single female reached the age of 5 years. Clear-cut habitats do not allow female M. vancouverensis to attain the same lifelong reproductive performance which their counterparts in natural habitats exhibit. I suggest that colonies in clearcuts are maintained by continual immigration of new females, and effectively function as demographic "sinks" (see Pulliam 1988).
M. vancouverensis is now confined to fewer than 15 mountains, mostly on south-central Vancouver Island (Bryant & Janz in press). One interpretation of the current distribution is based on the idea that historically-occupied habitats are no longer suitable because of climatic trends and associated vegetation changes (Nagorsen et al. in press). This hypothesis is the logical extension of Milko's (1984) work, who suggested that invasion of sub-alpine meadows by trees limits marmot habitat, or that marmots could be limited by food availability (Martell & Milko 1986). Under this interpretation, sites formerly occupied by marmots have changed in some qualitative way, and the species is naturally confined to a shrinking geographic region in which suitable climatic and vegetation conditions are found. Evidence for this view includes recent finds of prehistoric bones in at least 5 locations, all well outside the core area of current distribution (Nagorsen & Keddie 1994, Nagorsen et al. in press). The "climatic regulation" hypothesis is consistent with the views of Thomas (1994), who argued that many rare species track environmental conditions, becoming locally extinct where conditions are no longer suitable, and colonizing sites where conditions improve.
The second interpretation of marmot distribution is based on a metapopulation and source-sink perspec-tive (Bryant 1990). Under the "source-sink" hypothesis, historic marmot habitats are vacant because local extinctions have not been balanced by recolonizations. This could occur if the essential ingredient of meta-population structure (successful dispersal or "connec-tivity") has been altered by human activity. Evidence for metapopulation structure in M. vancouverensis includes observations of local population fluctuations, extinctions and colonizations. Evidence for reduced connectivity is equivocal; the idea that newly created but low-quality habitats adjacent to natural meadows impede dispersal is supported by genetic data (Bryant 1990) and spatial concentration of colonization events (Bryant & Janz in press), but contrary evidence show-ing long-distance dispersal in a modified landscape is also available (Bryant in press, Bryant & Janz in press).
Resolution of these competing hypotheses will not be easy. A fundamental problem is the absence of an experimental "control" for work on M. vancouverensis (there is no unlogged watershed in which marmot colo-nies occur, and in which "normal" levels of landscape connectivity can be measured). The climatic hypothesis is also directly untestable. Although methods exist with which to measure habitat stability (e.g., dendro-chronology), it would be impossible to attribute absence of marmots from particular areas to cause-effect relationships based on changing climate.
The question of why marmots no longer inhabit some areas is of fundamental conservation significance. If climatic changes have altered historic habitats, efforts to re-establish colonies will fail and there may be little that managers can do to enhance marmot populations. If, on the other hand, the problem is human-caused alteration of landscape connectivity, then transplants may be a simple and economical means of population enhancement (e.g., Ramousse et al. 1992), and prospects for eventual "recovery" of the species are probably excellent.
I am greatly indebted to L. Dyck, D. Milne, and J. Robin, who assisted in the field, and to the Vancouver Island Marmot Recovery Team, who provided unflagging support for research and management of M. vancouverensis. Funding for this study was provided by the B.C. Habitat Conservation Fund, B.C. Ministry of Environment, Lands and Parks, World Wildlife Fund (Canada), Cowichan Valley Field Naturalists Society, Forest Alliance of B.C., and Canadian Wildlife Service. MacMillan Bloedel Limited and Timber West allowed unrestricted access to private property. The Ecological Reserves Unit of B.C. Parks permitted me to trap marmots in the Haley Lake Ecological Reserve.
BRYANT A.A. In press. Reproduction and persistence of Vancouver Island Marmots
(Marmota vancouverensis) in natural and logged habitats. Canadian
Journal of Zoology.
BRYANT A.A. 1990. Genetic variability and minimum viable populations in the
Vancouver Island marmot (Marmota vancouverensis). M.E.Des. Thesis,
University of Calgary (Calgary, Alta.). 101 pp.
BRYANT A.A. & JANZ D.W. In press. Distribution and abundance of Vancouver
Island Marmots (Marmota vancouverensis). Canadian Journal of
Zoology.
CAUGHLEY G. 1977. Analysis of Vertebrate Populations. 1977. John Wiley
and Sons (New York, NY), 234 pp.
HEARD D.C. 1977. The Behavior of Vancouver Island Marmots (Marmota
vancouverensis). M.Sc. Thesis, University of British Columbia (Vancouver,
BC), 129 pp.
JANZ D. C., BLUMENSAAT N.K., DAWE B., HARPER S., LEIGH-SPENCER W. MUNRO &
D. NAGORSEN 1994. National Recovery Plan for the Vancouver Island Marmot.
Report No. 10, Recovery of Nationally Endangered Wildlife Committee, Ottawa.
MARTELL A.M. & R.J. MILKO 1986. Seasonal diets of Vancouver Island
marmots. Canadian Field-Naturalist, 100:241-245.
MILKO R.J. 1984. Vegetation and foraging ecology of the Vancouver Island
marmot (Marmota vancouverensis). M.Sc. Thesis, University of
Victoria (Victoria, BC), 127 pp.
MUNRO W.T., D.W. JANZ, V. HEINSALU & G.W. SMITH 1985. The Vancouver Island
Marmot: status and management plan. B.C. Ministry of Environment Wildlife
Bulletin B-39, (Victoria, BC), 24 pp.
NAGORSEN D.W. 1987. Marmota vancouverensis. Mammalian Species,
270:1-5.
NAGORSEN D.W., & KEDDIE G. 1994. Paleo-archeological remains of the
Vancouver Island marmot (Marmota vancouverensis); implications for an
endangered species. Abstract, American Society of Mammalogists, 75th
Anniversary Meeting, June 18-23, Washington, DC.
NAGORSEN D.W, G. KEDDIE & T. LUSZCZ. In press. Archaeological cave bones of
the Vancouver Island marmot. Occasional Paper #?, B.C. Ministry of
Environment, Lands and Parks (Victoria, BC).
PULLIAM H. R. 1988. Sources, sinks and population regulation. American
Naturalist, 132 : 652-661.
RAMOUSSE R., J.P. MARTINOT, & M. LE BERRE 1992. Twenty years of
re-introduction policy of alpine marmots from the national park of La Vanoise
(French Alps). In: Proceedings of the 1st International Conference on
Genus Marmota, Bassano, B., P. Durio, U. Gallo Orsi and E. Macchi
(editors), (Torino, IT), 171-177.
SWARTH H.S. 1911. Two new species of marmots from Northwestern America.
University of California Publications in Zoology, 7: 201-204.
THOMAS C.D. 1994. Extinction, colonization and metapopulations: environmental
tracking by rare species. Conservation Biology, 8:373-378.
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |