Mots-clés : Marmotte alpine, Marmota marmota, Réserve Naturelle Grande Sassière, coproscopies, bilans parasitaires, accroissement pondéral.
![]() Le suivi annuel de marmottes marquées de la Réserve Naturelle de
la Grande Sassière a permis d'obtenir les données suivantes sur
la parasitofaune de leur tube digestif et ses conséquences sur la
dynamique de leur population : 1. La cinétique des parasites a
été évaluée grâce à des coproscopies
périodiques durant la saison d'activité. Elle met en
évidence une forte variabilité temporelle des espèces
dominantes : Ctenotaenia marmotae, Citellina alpina,
Ascaris laevis et Eimeria sp. 2. L'accroissement de la masse
corporelle des animaux traités par un vermifuge anthelminthes a
été comparé à celui des animaux d'un groupe
témoin. Le but de cette étude est d'évaluer le rôle
joué par les parasites sur l'accumulation lipidique et le taux de survie
chez la marmotte alpine.
Le suivi annuel de marmottes marquées de la Réserve Naturelle de
la Grande Sassière a permis d'obtenir les données suivantes sur
la parasitofaune de leur tube digestif et ses conséquences sur la
dynamique de leur population : 1. La cinétique des parasites a
été évaluée grâce à des coproscopies
périodiques durant la saison d'activité. Elle met en
évidence une forte variabilité temporelle des espèces
dominantes : Ctenotaenia marmotae, Citellina alpina,
Ascaris laevis et Eimeria sp. 2. L'accroissement de la masse
corporelle des animaux traités par un vermifuge anthelminthes a
été comparé à celui des animaux d'un groupe
témoin. Le but de cette étude est d'évaluer le rôle
joué par les parasites sur l'accumulation lipidique et le taux de survie
chez la marmotte alpine.
Mots-clés : Marmotte alpine, Marmota marmota, Réserve Naturelle
Grande Sassière, coproscopies, bilans parasitaires, accroissement
pondéral.
The monitoring of marked marmots in La Grande Sassière Natural
Reserve during a year cycle allowed to get data on following topics about
parasitic fauna of their digestive tract and consequences on their population
dynamic: 1. The parasite kinetic was evaluated by use of periodic coproscopies
during the activity season. It shows a high variability of dominant species:
Ctenotaenia marmotae, Citellina alpina, Ascaris laevis and
Eimeria sp. 2. The body mass increase was compared between a treated set
(animals with antihelmintic vermifuge) and a control set. This study aims to
evaluate the role played by parasites on fat accumulation and winter survival
rate in alpine marmots.
Key-words: Alpine marmot, Marmota marmota, La Grande Sassière Nature
Reserve, coproscopies, parasite outcome, mass increase.

Chez la Marmotte alpine (Marmota marmota), le parasitisme peut atteindre des niveaux exceptionnels, en comparaison avec d'autres mammifères sauvages, comme l'ont montré de récentes études (Montagut & Prud'homme 1989 ; Sabatier 1989 ; Bassano 1992) : par exemple, la biomasse parasitaire pour les cestodes en particulier peut représenter jusqu'à 2% du poids corporel d'un individu. Il ne semble pas induire de symptôme clinique mais il peut être à l'origine de conséquences indirectes, particulièrement pour la dynamique de population.
Nous avons donc cherché à répondre à deux questions : 1. Comment l'important parasitisme digestif de la marmotte peut-il influencer la constitution de ses réserves graisseuses ? 2. Est-il susceptible de limiter ses chances de survie hivernale et d'influencer la régulation de ses populations ?
Au cours du printemps-été 1994, nous avons cherché à établir les cinétiques parasitaires de la marmotte sur un site et à comparer l'accroissement pondé-ral d'un lot d'animaux vermifugés à celui d'un lot d'animaux témoins, dans des conditions de vie naturelle.
En 1994, environ 150 marmottes ont été capturées sur le
site grâce à des <
Nous avons utilisé, toujours conjointement, le Praziquantel (DRONCIT
ND), un cestodicide spécifique (à la dose de 0,1
ml/kg) et l'Ivermectine (IVOMEC ND), vermifuge plus polyvalent
(à 0,25 ml/10 kg). Aucun de ces produits n'a d'action sur les
Coccidies.
Nous avons de plus essayé un traitement avec l'Arécoline,
vermifuge réputé pour ses capacités de purge
immédiate pour les cestodes chez les carnivores : il n'a eu aucun effet
chez la marmotte !
Le lot témoin a été constitué par des animaux
capturés au hasard sur les autres groupes sociaux du site ; il est
aussi constitué de dix-huit individus.
Le suivi de l'accroissement pondéral des animaux des deux lots a
été réalisé d'une part par pesée
systématique de tous les animaux capturés, d'autre part par
l'utilisation d'une méthode originale de pesée automatique des
animaux sur le terrain, sans manipulation. Nous avons pour cela disposé
sur le terrier principal des groupes étudiés une balance
électronique, connectée à un ordinateur de poche. Ce
système permet d'enregistrer l'heure et le poids indiqué par la
balance, pendant qu'un observateur à distance note l'identification de
l'animal, ses mouvements et l'heure de son passage sur le plateau. Le programme
réalisé spécifiquement pour cette étude fonctionne
en deux temps : il enregistre la tare toutes les dix secondes quand aucun
animal n'est présent (pour tenir compte des variations possibles de
cette tare, suite à des coups de vent par exemple), puis toutes les
secondes quand le poids est supérieur au seuil de 3 kg, donc quand un
animal est sur le plateau.
Le socle en bois et le plateau étaient laissés en permanence sur
le terrain ; le dispositif de pesée était cachés dans le
socle, juste avant les phases d'observation, de préférence
tôt le matin avant la sortie des premières marmottes. Pour inciter
les marmottes à monter sur la balance nous avons réalisé
plusieurs essais : un entonnoir en grillage, des appâts, des tubes
de marquage (photo 1).
Nous allons dans l'avenir chercher à améliorer le protocole en
utilisant un système de déclenchement de la pesée
à distance, pour éviter les manipulations journalières du
système et laisser aux marmottes le temps de s'adapter au
dérangement.
L'hypothèse d'une persistance hivernale et d'une réactivation au
printemps par des formes dormantes nous semble validée.
Le tableau 1 montre que 100% des groupes sont infestés du début
juillet jusqu'en fin septembre au moins. Ctenotaenia marmotae est
donc bien le parasite dominant de la communauté d'helminthes de la
marmotte (Manfredi et al. 1992).
Enfin, le suivi coproscopique montre que des oeufs sont aussi émis fin
avril-début mai. Ce résultat est en totale opposition avec les
données de la littérature : étant donné la longue
période prépatente de ce parasite, cette émission ne peut
pas être la conséquence d'une infestation de l'année. Nous
pouvons donc émettre deux hypothèses pour expliquer ce
phénomène : 1. Les fèces analysées étaient
en fait des prélèvements de la fin de l'année
précédente, bien conservées par le gel et la neige. 2. Des
parasites transhivernants survivent sous une forme particulière dans le
tractus digestif complète-ment atrophié et vide de la marmotte en
hibernation. Comme cela a été montré chez des oiseaux
migrateurs par Graber (comm. pers. Gevrey), ils pourraient subir un
phénomène de <
Mis à part ce phénomène à élucider par des
études ultérieures, les résultats obtenus à la
Grande Sassière sont très proches de ceux rapportés par la
littérature dans d'autres sites des Alpes.
Nous avons comparé l'évolution pondérale des animaux des
deux lots en calculant leur Gain Moyen Quotidien (GMQ) ; les résultats,
regroupés dans le tableau 5, montrent que la différence globale
n'est pas significative entre les deux lots. Par contre individuellement, nous
avons noté de très grandes variations dans le lot
vermifugé. Par exemple, l'animal dont le GMQ n'a été que
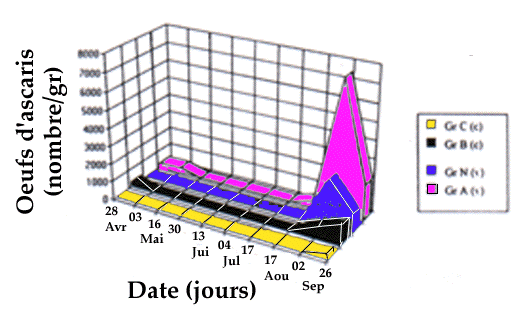
de 9g/j après vermifugation, montre une très forte
émission d'oeufs d'ascaris en août, précédant le pic
général montré par cette population. Cette charge en
ascaris peut tout à fait expliquer le retard de croissance
observé chez ce jeune de un an. Un autre résultat similaire
apparaît sur les figures 6 et 7. Nous remarquons que la dynamique
d'excrétion pour les groupes traités est sensiblement
modifiée par rapport à celle des groupes témoins. Pour les
oeufs d'ascaris, nous obtenons un pic élevé correspondant
exactement à la période prépatente de l'ascaris. Pour les
ookystes, nous obtenons une émission variable mais toujours plus
élevée que celle des groupes témoins en fin de saison.
Nous pouvons expliquer ce phénomène de deux façons :
* L'élimination des parasites adultes en juin aurait levé
l'inhibition des larves de stade 5 d'ascaris et favorisé leur maturation
massive.
* La vermifugation aurait perturbé profondément la
communauté d'helminthes de la marmotte telle que dé-crite par
Manfredi (1992). L'équilibre obtenu naturellement par
compétitions interspécifiques entre Cteno-taenia marmotae,
Ascaris laevis et Citellina alpina est détruit. Les
espèces restantes et les formes infestantes envahissent alors
l'écosystème digestif laissé libre.
Notre étude confirme l'importance du parasitisme intestinal chez la
Marmotte alpine. Malgré l'absence de symptôme clinique, ce
phénomène joue probablement un rôle
prépondérant dans la régulation de sa survie hivernale.
Contrairement aux ongulés pour lesquels les périodes
d'infestations sont temporaires, il semble y avoir une forte pression
parasitaire durant toute la période d'activité des marmottes.
Un autre point original de l'étude concerne une éventuelle
persistance parasitaire durant l'hibernation : les Ctenotaenia marmotae
sont-ils capables de <
Enfin, l'intervention humaine montre ici ses limites, par des perturbations
inattendues et prouve combien nous devons être prudents dans la gestion
de la faune sauvage.
Tableau 3. Résultats coproscopiques pour M. 1 an ndeg.5, groupe N.
Figure 1. Eimeria cycle
Figure 3. Ascaris laevis cycle
Figure 4. Dynamic of Ascarid eggs shedding
Figure 5. Ctenoteania cycle
*****
Several authors have described the influence of parasitism on their
host-population, either on physiological events (reproduction, food
performance), or behaviour (social status in mice: Freeland 1981; young
dispersal in swallows: Brown & Bromberger-Brown 1992; habitat use in
reindeer: Folstad et al. 1991) or in dynamic of populations (Barbault
1993).
For alpine marmots (Marmota marmota), parasitism appears to reach
exceptional levels, when compared to other mammals, according to recent studies
(Montagut & Prud'homme 1984, Sabatier 1989, Bassano 1992): for example, the
biomass of intestinal helminths represents up to 2% of the total weight of
parasitized marmots. It doesn't seem to induce patho-logical symptoms, but it
may lead to indirect consequences, especially for dynamic of populations.
So the questions we wanted to answer were:
* How does the important parasitism of marmots interfere with the
constitution of their fat reserves?
* Does it interfere with their winter survival probability and with the
regulation of their dynamic of population ?
So, during spring and summer 1994, we have been starting a study in National
Park of Vanoise, with two directions:
* Setting the parasite kinetics of marmots of this area.
* Comparing the weight increase of animals treated with vermifuge, with the
weight increase of control animals, under wild life conditions.
During 1994, about 150 marmots have been trapped with wire-netting boxes and
lured with apple or pop-corn baits. Trapped marmots were marked with coloured
collar and eartags, so that each individual could be identified from a fair
distance.
These faeces were analysed in the Veterinary Laboratory of Savoy. We used a
floating method with iodomercurate of potassium (density = 1.44), then counting
eggs and larvae of helminths and oocysts of Coccidia, with Mac Master
cells.
We injected at the same time Praziquantel, a specific cestodicid
(DRONCIT ND, 0.1 ml/kg intra-muscular) and Ivermectine, a
polyvalent product (IVOMEC ND, 0.25 ml/10 kg under the skin).
They were not efficient on coccidies. We also tried a treatment with Arecoline,
which is said to violently purge the cestods of carnivores, but it didn't work
at all on marmots!
The control set was constituted of 18 animals caught at random on the other
social groups.
The weight increase survey of the two sets has been carried out with two
methods: trapped marmots were systematically weighted with manual scales and we
tried to set an original method of automatic weighting, working on site without
disturbing the animals. This method consisted of an electronic scale connected
to a pocket-computer able to record the weight/time data, when an observer
noticed identifi-cation and movement of animals and time. All these instruments
were hidden in a wooden box, buried just in front of the marmot burrow. The
specific computer program of this study allowed for the tare every ten second,
in order to appreciate the interferences due to wind and dust-carrying, and the
weight was recorded every second as soon as the total weight exceeded 3 kg, so
when a marmot was on the scale.
The wooden box was permanently on the burrow but the weight system was
hidden every day just before the observations. The standing of marmots on the
scale-cover was helped by different expedients: wire-netting corral, baits....
(Photo 1).
One improvement will be to set a distance transmitter to switch on and off
the recording of measurement in order to avoid human interference during the
whole week, period needed for marmots to get used to stroll of the
scale-cover.
We noticed that these coccidia were constantly present in all studied
groups, even after hibernation and that the highest shedding occurred in spring
(figure 2), as described by Bassano et al. (1992). Our results are in
accordance with the hypothesis of winter persistence of coccidial infestation
and with a reactivation from "dormant-stage" at this time.
On the other hand, our coproscopic survey showed that eggs were already shed
in the end of April and in early May. This result is in opposition with the
data found in literature: according to the indispensable prepatent period, this
shedding can't be the consequence of early spring infestation. Two hypothesis
can be given to explain this fact:
* The faeces were issued from the previous year and were kept through
winter, under the protecting cover of snow.
* They were really overwintering worms living under an original state.
Several authors (Blanchard & Blatin 1907, Couturier 1963, Bassano et
al. 1992, for example) have described an intestinal vacuity in hibernating
marmots, digestive tract is atrophied and intestinal lumen is absolutely
virtual. So the overwintering form might be under a microscopic stage such as
described in some migrating birds (by Graber, pers. com. Gevrey), under
unfavourable conditions, only the scolex of cestods are remaining in intestine
called "destrobilisation" phenomenon).
Except with this phenomenon that needs further investigations, our results
confirm other studies in the Alps.
We could then think that the treatment has been efficient, at least
temporarily. Regular survey of social group N, the members of which were all
treated with vermifuge, agreed with this result (table 4).
It shows the lack of nematode eggs and of cestode eggs and proglottis in the
month following the vermifugation, but a persistence of coccidia infestation,
which agrees with the theoretical antihelminthic activity spectrum of the two
products.
New shedding of eggs was detected again starting in the middle of august:
considering the prepatent period, it shows that the infestation pressure keeps
on being on a high level during summer, and as a consequence, that
vermifugation only brings a momentary effect.
Then, we analysed the daily weight-increase (GMQ) of marmots under
vermifuge, compared to control set, in table 5. The global difference between
the two sets was not significant. But among the three animals under vermifuge,
variations were observed. The case of the animal having a low increase after
vermifuge is interesting to describe: the coproscopic survey showed a high
shedding of ascarid eggs in August, occurring before the peak of marmot
population.
Another similar results is shown in figures 6 and 7: we noticed that the
dynamic of shedding in groups under vermifuge is modified, compared to control
groups. For ascarid eggs, a higher peak was connected to the end of a prepatent
period starting at vermifugation. For oocysts, we detected a varying shedding,
at an upper level compared to control groups.
The explanation may be the following:
* The elimination of parasite adults might have disturbed the equilibrium
and change the intestinal parasitic fauna. For example the disappearance of
ascaris adults improves the inhibition of fifth stage larvae and improves their
mass-maturation into aggressive young adults.
* Manfredi (1992) expressed the concept of helminth community of marmot,
with interspecific relationships between Ctenotaenia marmotae, Ascaris
laevis and Citellina alpina. When inducing a disturbance in the
interspecific competition in equilibrium with the host (for example, with the
disappearance of the dominant species), the remaining species expand in the
newly free ecosystem.
Our study confirmed the importance of intestinal parasitism in Alpine
marmots. Although there is a lack of clinical manifestations this phenomenon
may play a prominent part in regulation of their hibernation survival
probabilities.
Contrarily to ungulates where infestation periods are temporary, a high
pressure seems to persist during the whole activity-period of marmots.
Another original point is the question of hibernation infestation: is the
Ctenotaenia able to "destrobilisation" and quick weight increase in
early spring, showing there a surprising coadaptation to hibernation? Further
studies will be necessary to check this hypothesis.
At least the human trends to interfere in natural ecosystems demonstrate
here their limits and how we must be cautious in the management of
wildlife.
A suivre
B.
ANALYSES COPROSCOPIQUES, POUR LES CINÉTIQUES PARASITAIRES.
Nous avons prélevé des fèces de marmottes, environ tous
les quinze jours, soit au hasard sur les latrines des groupes
étudiés, soit individuellement dans les cages de capture. Les
analyses ont été réalisées au Laboratoire
Vétérinaire de la Savoie, avec la méthode de coproscopie
par flottaison à l'iodo-mercurate de potassium (d = 1,44) en utilisant
une lame Mac-Master et une lame de flottaison pour compter les oeufs
d'helminthes et les ookystes de Coccidies, .
C.
CAMPAGNE DE VERMIFUGATION ET ACCROISSEMENT PONDÉRAL DES ANIMAUX
Dix-huit marmottes, appartenant à deux groupes sociaux, ont
été vermifugées entre fin juin et début juillet,
période principale d'infestation décrite par Sabatier (1989).
RÉSULTATS
ET DISCUSSION
A.
PARASITES RENCONTRÉS
Les espèces parasites dominantes, que nous avons rencontrées,
sont Ctenotaenia marmotae, Citellina alpina, Ascaris
laevis et Eimeria sp., comme le montre le tableau 1.
1.
Coccidies du genre Eimeria
Ce sont des protozoaires, parasites intra-cellulaires. Leur cycle
évolutif est monoxène (figure 1). La contamination de nouveaux
hôtes se fait grâce à la dissémination des ookystes,
très résistants dans le milieu extérieur. Sur le site
étudié, les coccidies sont omniprésentes sur toute la
période et dans tous les groupes. Le pic d'émission des ookystes
au printemps est très net (figure 2), comme l'ont montré Bassano
et al. (1992) sur d'autres zones.
2.
Citellina alpina
C'est un nématode de la famille des Oxyuridés. Comme les autres
membres de cette famille, il présente une très forte
spécificité d'hôte : Citellina alpina n'est
présent que chez Marmota marmota. Sabatier (1989) parle d'un
phénomène de coévolution phylogénétique
stricte entre les genres Citellina et Marmota. Comme
décrit dans la littérature, la période d'émission
des oeufs est très courte et la méthode coproscopique ne semble
pas très fiable pour ce nématode : nous n'avons
détecté Citellina dans les fèces que fin mai /
début juin, alors que les études par autopsies montrent que c'est
un des parasites majeurs de la marmotte.
3.
Ascaris laevis
Ce parasite est aussi très spécifique de la marmotte ; son cycle
évolutif est monoxène (figure 3), mais il nécessite une
période de maturation des oeufs dans le milieu extérieur, de
durée inconnue. Par comparaison avec les ascaris de Marmota
sibirica, décrits par Mozgovoï (1953), la période
prépatente est estimée à 40 jours. Les résultats
coproscopiques, bien corrélés avec la charge parasitaire,
montrent un pic d'émission élevé en septembre (figure
4).
4.
Ctenotaenia marmotae
Ce cestode Anoplocéphale présente un cycle évolutif
dixène (figure 5). Les adultes vivent dans le tube digestif de
l'hôte définitif. Leur capacité à absorber les
nutriments directement à travers leur surface corporelle (Smyth 1969),
les rend responsables d'une forte spoliation de nourriture pour leur
hôte. Les proglottis entiers sont en général émis
dans les fèces, donc nous ne pouvons détecter les oeufs en
coproscopie que lorsque ces segments sont déchirés ou
écrasés. L'hôte intermédiaire est un acarien
Oribate, dans lequel la maturation des oeufs demande au moins 4 mois (Ebermann
1976).
B.
CONSÉQUENCES DU TRAITEMENT SUR L'ACCROISSEMENT PONDÉRAL
L'efficacité du traitement a été vérifiée
par coproscopies avant et après vermifugation. Ainsi, les tableaux 2 et
3 montrent une élimination instantanée pour les deux individus F
et M des Ascaris et des Cestodes. Le suivi régulier du groupe N dont
tous les individus ont été traités corrobore ce
résultat (tableau 4). Il montre l'absence d'oeufs de nématodes et
de cestodes dans le mois qui a suivi la vermifugation (analyses du 24 juillet),
mais il y a persistance de l'infestation coccidienne, ce qui concorde avec le
spectre d'activité des produits utilisés. Puis à partir de
la mi-août, les coproscopies sont de nouveau positives : compte tenu de
la période prépatente, cela montre que la pression d'infestation
s'est toujours maintenue à un niveau important durant
l'été.
CONCLUSION
Table 1. Qualitative results of copromicroscopic survey. Caractéristiques des prélévements % des groupes positifs pour :
Date Nombre de groupes Ookystes (Eimeria) Oeufs de Cestodes (Ctenotaenia) Oeufs d'Ascaris (Ascaris laevis) Oeuf d'Oxyures (Citellina)
28 avr 3 100% 67%(2/3) 100%(3/3) 0
03 mai 6 100% 50%(3/6) 50%(3/6) 0
11 mai 4 100% 50%(2/4) 25%(1/4) 0
16 mai 4 100% 0(0/4) 25%(1/4) 0
30 mai 6 100% 17%(1/6) 0%(0/6) 0
13 jui 5 100% 60%(3/5) 60%(3/5) 20%(1/5)
08 jui 4 100% 100%(4/4) 0(0/4) 25%(1/4)
24 jui 4 100% 100%(4/4) 25%(1/4) 0
17 aou 3 100% 100%(3/3) 100%(3/3) 0
02 sep 3 100% 100%(3/3) 100%(3/3) 0
26 sep 4 100% 100%(4/4) 100%(4/4) 0
06 oct 4 100% 100%(4/4) 100%(4/4) 0
Table 2. Coproscopic results of 1 year F ndeg.1, group N.Date Ascaris Cestodes Oxyures Coccidies
13-06 135/g présence 0 2.880/g
08-07 0 0 0 13.440/g
Table 3 : Coproscopic results of 1 year M ndeg.5, group N.Date Ascaris Cestodes Oxyures Coccidies
17-06 0 présence 0 120.800 /g
20-08 110/g 0 0 1.800/g
Table 4. Coproscopic results of group N, after first treatment.Date Ascaris Cestodes Oxyures Coccidies
13-06 0 0 0 16.400/ g
08-07 0 présence 0 8.400/g
24-07 0 0 0 12.600/g
17-08 45/g présence 0 240/g
02-09 2.560/g 0 0 500/g
Gr Identité des individus GMQ global GMQ début de saison GMQ postV ou fin de saison
N F.1 an ndeg.2 34g/ j ? 34g/j
A M.1 an ndeg.2 14g/ j 20g/j 9g/j
C M.1 an ndeg.1 27g/ j 21g/j 29g/j
Moyenne des V 25g/ j 20g/j 24g/j
C F.1 an ndeg.2 25g/ j 25g/j 25g/j
I F.1 an ndeg.1 25g/ j 25g/j 25g/j
I F.1 an ndeg.2 25g/ j 25g/j 25g/j
I F.1 an ndeg.3 28g/ j ? ?
E M.1 an ndeg.1 19g/ j 19g/j ?
E M.1 an ndeg.2 21g/ j 21g/j ?
E F.1 an ndeg.3 17g/ j 17g/j ?
Moyenne des non-V 23g/ j 21g/j 25g/j
Table 6. Coproscopic results of 1 year M ndeg.2, group A.Date Ascaris Cestodes Oxyures Coccidies
16-06 0 présence 0 5.340/g
24-08 10.440/g présence 0 113.000/g
Photo 1. Automatic scale in the field
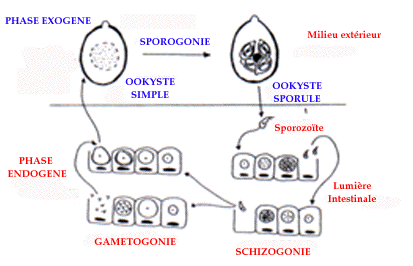
Figure 2. Dynamic of oocysts shedding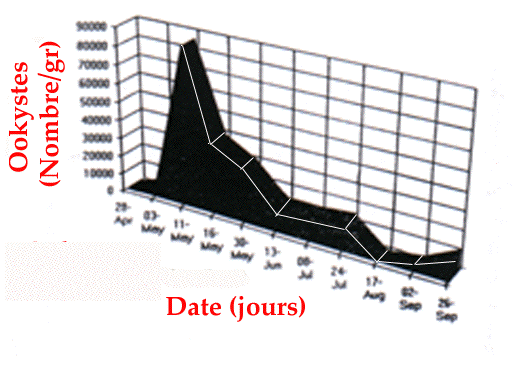
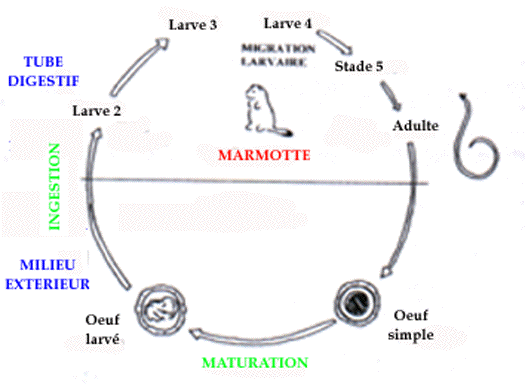

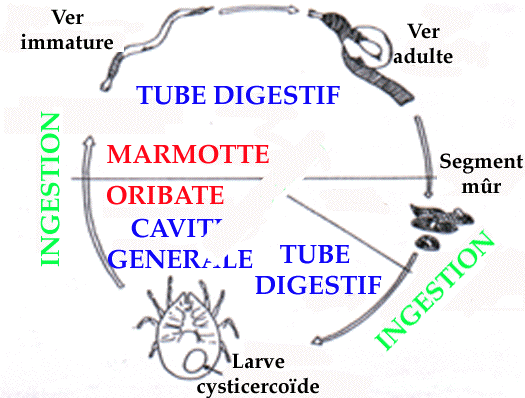
Figure 6. Dynamic of oocysts shedding in traited (v) and control groups (t).
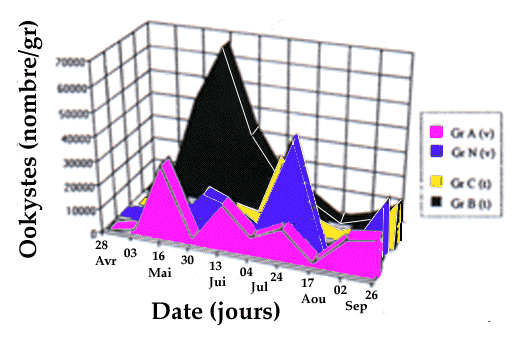
Figure 7. Dynamic of ascarid eggs shedding in traited (v) and control
groups (c).
INTRODUCTION
The concept of parasitism has been perceptibly evolving during the last ten
years, according to the recent ecology theories. Formerly it was said to be an
abnormal phenomenon, damaging the health of domestic cattle; nowadays it is
considered as an important factor of regulation of wild ecosystems, a trophic
interspecific relationship overspread in animal kingdom.
MATERIAL
AND METHODS
A.
STUDY AREA, CAPTURES AND ANIMAL IDENTIFICATIONS
The study area is set in La Grande Sassière Natural Reserve,
north-east of Vanoise National Park, near the Italian border.
B.
COPROMICROSCOPIC ANALYSIS, FOR PARASITIC KINETICS
Every fifteen days, marmots dropping were taken either at random from the
"latrines" of social groups under study, or from some identified animals during
captures.
Eighteen marmots, belonging to two social groups, were treated with
vermifuge by the end of June and beginning of July, periods which are likely to
give the main infestations (Sabatier 1989).
C.
VERMIFUGE CAMPAIGN AND WEIGHT INCREASE
RESULTS
AND DISCUSSION
A.
PARASITES FOUND
The dominant species of parasites we have found are Ctenoteania
marmotae, Citellina alpina, Ascaris laevis and Eimeria sp., as shown
in table 1.
1.
Coccidia of Eimeria genus
They are intra cellular protozoan parasites. Their cycle is monoxen (figure
1). The contamination of other hosts is obtained by excretion of oocysts which
are very resistant in the field.
2.
Citellina alpina
This nematode belongs to the family of Oxyuridae. It shows, as the
other members of this parasite-family, a strong host-specificity: Citellina
alpina is only found in Marmota marmota. Sabatier (1989) speaks of a
strict phylogenetic co-evolution phenomenon between these two genus. As found
in the literature, egg-shedding is very short, and it may explain the low
sensibility of coprologic method for this nematode. Actually, we found
Citellina alpina only in late June/early July in La Grande
Sassière, whereas autopsy survey use to show Citellina as a
dominant species for marmots.
3.
Ascaris laevis
This nematode is also specific to marmots in our country. Its cycle is
monoxen (figure 3), but needs a maturation period in external environment.
Referring to the Ascaris of Marmota sibirica (Mozgovoï
1953), the prepatent period is nearly 40 days. The coprologic results
(considered as well correlated with parasitic load) showed a high peak in
September (figure 4).
This cestode needs two hosts (figure 5): the adult-form lives in digestive
tract of marmot, where it feeds through its whole body-surface (Smyth 1969): so
it might have a real despoiling effect on marmots. The mature proglottis are
shed in full, so that the coprology only detects eggs when these segments are
torn. The intermediate host is an Oribatid; the total evolution period inside
this host needs nearly 4 months according to Ebermann (1976). Table 1 shows
that 100% of groups were infested from the beginning of July to October: this
confirms the dominance of this parasite in marmot helminth community (Manfredi
et al. 1992).
4.
Ctenotaenia marmotae
B.
EFFECT OF ANTIHELMINTHIC ADMINISTRATION ON WEIGHT INCREASE
Efficiency of treatment were controlled with coproscopic survey before and
after antihelminthic administration, for two animals (tables 2 and 3).
CONCLUSION
BIBLIOGRAPHIE/REFERENCES
BARBAULT R. 1993. Écologie générale. Structure
et fonctionnement de la biosphère. Masson collection
Abrégés, 2ème édition, 269pp.
BASSANO B., SABATIER B., ROSSI L. & MACCHI E. 1992. Parasitic fauna on the digestive tract of Marmota marmota in the western Alps. In Proc. 1 st
International Symp. on Alpine Marmots, Bassano B., Durio P., Gallo Orsi U.,
Macchi. Eds, 13-24.
BLANCHARD R. & BLATIN M. 1907. Immunité de la marmotte en
hibernation à l'égard des maladies parasitaires. Arch. Parasitol., 11, (3): 362-378.
BROWN C.R. & BROMBERGER-BROWN M. 1992. Ectoparasitism as a cause of natal dispersal in cliff swallows. Ecology, 67: 1206-1218.
COUTURIER M.J.A. 1963. Contribution à l'étude du sommeil hibernal chez la marmotte des Alpes, Marmota marmota (L.) 1758. Mammalia, 27(4): 456-482.
EBERMANN E. 1976. Oribatids (Oribatei, Acari) as intermediate
hosts of Ctenotaenia marmotae (Frölich. 1802) a tapewom parasitc in
marmots. Z. für Parasitenkunde, 50: (3) 303-3 12. (ln german).
FOLSTAD L., NILSSEN A.C., HALVORSEN O. & ANDERSEN J. 1991. Parasite avoidance: the post-calving migrations in Rangirfer ? Can. J. Zool., 69:
2423-2429.
FREELAND W.J. 1981. Parasitism and behavioral dominance among male mice.
Science, 213: 46l-462.
MANFREDI M.T, ZANIN E. & RIZZOLI A.P. 1992. Helminth community on alpine marmots. In Proc. 1st Symposium International on Alpine
Marmots, Bassano B., Durio P., Gallo Orsi U., Macchi E., Eds: 203-207.
MONTAGUT G. & PRUD'HOMME C. 1989. Helminthes parasites de la marmotte dans le massif de la Vanoise, observation d'une infesfation par
Capillaria hepatica. Tr. Sci. Parc Nation. Vanoise, l 7:
229-234.
MOZGOVOï A.A. 1953. Ascaridata of Animals and Man and the diseases caused by them. Part 1, The Academy of Sciences of USSRR Ed, Moscow, 390pp.
SABATIER B. 1989. Les parasites de la marmotte alpine: étude dans les
Alpes françaises et synthèse bibliographique. Thèse
Ecole Nationale Vétérinaire de Lyon, 178pp.
SMYTH J.D. 1969. The physiology of Cestodes. Oliver and Boyd LTD,
Edinburgh.
Forward![]()
Retour à Documents scientifiques du réseau
Back to Scientific publications of the network sommaire/contents ![]()