![]()
Les différences de taille et de longueur condilo-basale du crâne
entre les marmottes eurasiennes et nord-américaines sont à peine
perceptibles. Des espèces et des sous-espèces de toutes tailles
se trouvent sur les deux continents. Aucune augmentation de la taille et de la
longueur condilo-basilaire n'est observée dans les parties
septentrionales et les hautes montagnes de leur distribution. Les durées
d'activité estivale et d'hibernation varient d'une espèce
à l'autre ; la période d'hibernation est plus longue dans les
habitats à "été court". Des adaptations variées
rendent possible la vie de l'animal sous des conditions extrêmes avec une
période réduite d'activité estivale.
Mots-clés : Marmottes, taille corporelle, cycle annuel,
adaptations.
Difference in body size and condilobasal length of the skull between the
marmots of Eurasia and North America is hardly noticeable. Small and large
species and subspecies are found on both continents. No increase in the body
size and condilobasal length are observed in the northern parts of the range
and in the high mountains. The proportion of the duration of above-ground
activity and hibernation vary in different marmot species; the hibernation
period is prolonged in the habitats with "short summer". Various adaptations
make possible for the animals to live under extreme conditions and reduced
period of the above-ground activity.
Key-words: Marmots, body size, annual cycle, adaptations

Les marmottes (Marmota), largement distribuées dans la région Holarctique; occupent presque toutes les zones naturelles, des côtes de la mer arctique aux hautes montagnes d'Asie Centrale. Du fait de l'étendue de leur aire de distribution, elles pourraient sembler présenter une plasticité élevée pour l'affouragement, l'activité et le choix de l'habitat. En réalité, ces rongeurs coloniaux et hibernants sont des herbivores spécialisés au comporte-ment typique des habitants des espaces ouverts. Ils ont besoin non seulement d'une bonne vue de leur voisi-nage, mais aussi d'une épaisse couche de sol meuble pour creuser des terriers d'hiver.
Les périodes d'estivation et d'hibernation varient fortement selon les membres du genre. Chez la marmotte américaine M. monax, la période d'hibernation est très courte (2-3 mois) et même parfois absente, alors que chez M. camtschatica elle dure jusqu'à 8-9 mois. Ces variations s'expliquent par les conditions de vie des marmottes. La dépendance de l'intensité des pertes de chaleur à la taille corporelle des animaux, mise en évidence depuis longtemps, est moindre chez les animaux de grande taille (Morrison 1960, cit. Kalabukhov, 1985). Panteleev (1983) a noté qu'une perte thermique importante entraîne la sélection des individus à métabolisme élevé. Ainsi, l'adaptation à la sévérité des conditions de vie devrait s'accompagner d'une augmentation de la taille corporelle et de la masse des marmottes.
Pour la première fois nous comparons la taille corporelle de différentes espèces de marmottes en pre-nant en compte leur cycle annuel et leurs adaptations aux conditions particulières de vie pour tester les liens entre énergétique et taille des animaux. La majeure par-tie de la prairie Nord Américaine est occupée par des marmottes relativement anciennes du groupe Marmota (Gromov et al. 1965, Hoffman 1968). Les formes mo-dernes du groupe bobac, apparues à la fin du Pléisto-cène et à l'Holocène, l'emportent en Eurasie (Gromov et al. 1965, Bibikov 1967). Des formes plus anciennes oc-cupent les Alpes et les Carpates (M. marmota) et l'Asie Centrale (M. caudata et autres).
La comparaison des marmottes selon leur taille, critère morphologique le plus souvent utilisé, et leur masse (Fig. 1A) est réalisée pour la première fois com-me pour la longueur condilobasale du crâne (Fig. 1B), pour laquelle les erreurs de mesure semblent minimales. Les mesures des clés classiques (Howell 1915, Ognev 1947, Gromov et al. 1965, Lee & Funderburg 1982, Nowak 1991), et celles prélevées sur les espèces eura-siennes par les zoologistes (Petrov 1960, Tkachenko 1962) ont été utilisées. Les échantillons et les techniques de mesure ne sont naturellement pas iden-tiques dans les différentes études. J'ai parfois accordé une préférence aux données de certains auteurs. Les valeurs moyennes de chacune des espèces considérées ont été calculées à partir d'au moins 10 mesures. Dans certains cas, les données de différents auteurs ne sont pas comparables, il y a donc des problèmes de fiabilité, particulièrement pour la taille et la masse. Des marmottes de petite taille (M. menzbieri, M. flaviventris, M. broweri et M. camtschatica, surtout les sous-espèces Yakoutes et Barguzin) comme celles de grande taille (M. bobac, M. himalayana and M. caligata) se trouvent à la fois en Asie et en Amérique (Fig. 1A). La compa-raison de la longueur condilobasilaire confirme l'exis-tence de marmottes de différentes tailles et d'une transi-tion graduelle des animaux de petite à ceux de grande taille sur les deux continents.
Une attention spéciale a été accordée à l'exis-tence de différences géographiques et populationnelles de la taille de marmottes considérées comme apparte-nant à une même espèce. Les tailles de la marmotte à ventre jaune (M. flaviventris) varient d'une fois et demie entre les diverses parties de son aire de répartition et les différentes altitudes. Ces différences sont de l'ordre de deux fois chez les sous-espèces de la marmotte à tête noire (M. camtschatica). Suivant les populations, chez l'américaine M. flaviventris, les animaux peuvent être 1,5 fois plus petits et même plus de 1,5 fois plus légers que les marmottes du groupe caligata. Le même phénomène est observé entre les deux espèces paléarc-tiques, la marmotte de Menzbier et la bobac.
En ce qui concerne la géographie et l'écologie, je ne connais aucune tentative de comparaison entre les marmottes d'Asie et d'Amérique. Les zoologistes de l'Ouest ignorent presque la biologie des espèces eura-siennes, et nos spécialistes n'ont pas discuté de ce pro-blème non plus. C'est la raison probable de l'absence de toute discussion sur l'origine du large recouvrement des aires de répartition des espèces en Amérique et sur l'allopatrie en Europe et en Asie. Malgré des différences dans la biologie des marmottes, la similarité de certains détails, comme par exemple pour l'affouragement, l'activité, la structure de la population ou le compor-tement, est évidente. Il est remarquable que l'étendue des caractéristiques morphologiques et écologiques des individus d'une même espèce mais vivant dans des milieux ou des altitudes différents est parfois plus importante que celles entre les différentes espèces européennes et nord-américaines.
Première
étape.
Préparation à l'hibernation, déplacement
vers le terrier d'hibernation puis son occupation, dure environ 2,5 mois.
L'endormissement complet, chez les sous-espèces de bobac d'Europe et de
Mongolie, comme chez M. caligata et la marmotte du Tien Shan est plutôt
long et dure 2 à 3 semaines pour la même colonie (Ryabov 1948,
Barash 1976). Les membres d'une famille se concentrent d'abord à
proximité du terrier d'hivernage. La femelle reproductrice et ses jeunes
sont les derniers à cesser leur activité. Ces changements
comportementaux et la cessation des sorties des animaux sont en
corrélation avec les changements physiologiques des organismes. La
température corporelle chute la nuit à 28-32deg.C ; la
consommation d'oxygène décroît et d'autres modifications
physiologiques se produisent en même temps (Bliznetzov 1966, Vasilyev
1989). Habituellement la température interne de l'hiber-naculum varie de
0deg. à 5deg.C dans la zone de permafrost et en altitude. Après
la disparition des femelles et des jeunes à l'intérieur du
terrier hivernal, un adulte (?) en obstrue l'orifice, en réalisant un
bouchon, les autres ouvertures du terrier ayant été
scellées auparavant. Les premiers déterrages réussis par
Ryabov (1948) en Zabaikalie ont permis la mesure, pour la première fois,
de la température de la litière garnissant l'hibernaculum. Les
mesures télémétriques menées durant l'hibernation
chez la marmotte alpine (Arnold et al. 1991) ont confir-mé les
hypothèses de Ryabov concernant la chute graduelle de la
température dans l'hibernaculum, la présence d'animaux endormis
et les modifications physiologiques des animaux. A l'intérieur de
l'hiberna-culum bouché, l'obscurité est totale, la
température basse, l'humidité et la teneur en gaz carbonique
élevées, la teneur en oxygène faible. Selon la
théorie de l'autonarcose (Dubois 1896), l'augmentation de la teneur en
gaz carbonique dans l'hibernaculum entraîne le démarrage de la
torpeur (Bibikov 1967, p.72).
Il faut noter que la torpeur profonde débute fin octobre ou même
en décembre, chez M. baibacina et M. sibirica en
captivité, soit bien plus tard qu'en conditions naturelles. Ce passage
graduel (d'octobre à novembre) à l'état de torpeur
profonde est confirmé chez M. camt-schatica qui devient torpide
1,5 à 2 mois plus tard en captivité qu'en conditions naturelles.
En novembre leur température corporelle chute, la durée des
périodes de torpeur s'accroît alors que les intervalles les
séparant deviennent plus courts (Vasilyev 1989). Les expériences
menées avec M. bobac montrent, aussi, des changements (Rymalov
& Maximov 1992).
Deuxième
étape.
Elle dure 2 à 3 mois, fin no-vembre jusqu'à fin
février (Fig. 2.2). La plupart des animaux sont torpides et un petit
nombre d'entre eux est simplement assoupi. L'étude, sept années
consécutives, du régime de température et de la teneur en
oxygène de six terriers d'hivernage (Arnold et al. 1991) a montré
que 2,5 mois avant le démarrage de la torpeur profonde, la
température d'un hibernaculum à 1-3 m de profondeur chutait
brutalement (de 12deg. à 5deg.C) et que par la suite ce processus
devenait graduel (environ 1deg.C par mois).
Les arrêts réguliers de la torpeur au cours de l'hibernation,
phénomène très important, ont été
découverts pour la première fois chez l'écureuil terrestre
(Citellus lateralis ; Pengelley & Fischer 1961). Leur existence chez
la marmotte a été postulée par Berendyaeva et al.
(1967) après avoir mis en évidence les fluctuations de la
température dans des hibernacula, situés à 3400 m
d'altitude, de la marmotte du Tien Shan grâce à un
téléthermomètre AM-2M. Selon les auteurs, les
augmentations de température dépendent des éveils (tous
les dix jours) des marmottes hibernantes. Après décembre-janvier,
les périodes de torpeur de la mar-motte à tête noire
s'allongent et durent de 15 à 16,5 jours pour les femelles adultes, 17
j. pour les un-an et 20,5-22j. pour les marmottons (Vasilyev 1989).
3ème
étape.
Cette période de l'éveil final, de l'accouplement
et de l'abandon des terriers d'hiver est plus courte (mars ou avril-mai)
lorsque l'été est court (Fig. 2.3). Elle se caractérise
par des éveils périodiques plus fréquents et un nombre
plus élevé d'animaux som-nolents. Il reste une inconnue, comment
la ventilation de la chambre d'habitation est-elle réalisée lors
de la présence constante d'au moins une ou deux marmottes reproductrices
en activité ou simplement assoupies.
Le problème des dépenses énergétiques durant le
cycle annuel doit être discuté de façon plus
détaillé. La quantité de graisse sous-cutanée et
périviscérale cons-titue de 20 à 30% de la masse
corporelle à la veille de l'hibernation chez les marmottes eurasiennes
étudiées. Elle atteint 35% chez la marmotte à tête
noire de Yakoutie, valeur légèrement plus élevée
que celle de toutes les autres marmottes étudiées. Non seulement
la quantité de graisse, mais aussi les différences de sa
composition sont de grande importance. Sa composition en acide gras est
incomplètement connue, bien que l'on sache que l'acide gras
linoléique est plus abondant dans la graisse des sousliks de montagne
que dans celle des sousliks de plaine (Kalabukhov 1985). Les constantes
physico-chimiques de la graisse sont aussi très variables (Bliznetzov
1966).
Notre connaissance de la dynamique des dé-penses énergétiques au cours des étapes de l'hibernation est limitée. Chez M. monax, la perte totale de masse due à ces dépenses est de 1245 g , dont 41,6% sont perdues dans l'hibernaculum avant le démarrage de l'hibernation, 16,9% au cours de l'hibernation elle-même et 41,6% pendant l'éveil, mais avant le début de la période d'affouragement actif en mars (Bailey & Davis 1965). Au cours de la période d'activité, les marmottes eura-siennes, privées de nourriture, perdent 3,76% de leur masse corporelle par jour. Ces valeurs sont comprises entre 0,24 et 0,35% chez les rongeurs somnolents et seulement entre 0,02 et 0,12% chez les animaux tor-pides. Dans nos expériences, les marmottes du Tien Shan perdaient en moyenne 0,09 à 0,18% de leur masse corporelle de départ, ce qui correspond à une perte de masse absolue de 1,0 à 4,2 g (Bibikov 1967). Les dé-penses énergétiques chez cette marmotte sont plus im-portantes lorsque les animaux sont placés dans des conditions variables et relativement élevées de tempé-rature ambiante (8+/-3deg.C) que lorsqu'ils hibernent à une température constante (5deg.C ; Tab. 1).
Dans nos expériences nous avons porté l'atten-tion sur l'adaptabilité de l'hibernation continue et les dé-penses énergétiques plus réduites des jeunes. Nous avons supposé que les adultes les plus gras s'éveillent plus souvent et réchauffent l'hibernaculum facilitant ainsi le succès de l'hibernation des jeunes. Nos hypo-thèses ont été confirmées par une série d'études menées en Bavière (Arnold 1990, Arnold et al. 1991).
Les adaptations aux conditions extrêmes sont liées à la complexité et à la meilleure isolation des terriers d'hivernage : position favorable du terrier dans le relief, grande profondeur, épaisse litière, nombreux bouchons de terre (bouchons additionnels d'herbe dans les terriers des marmottes Yakoutes) et même une butte au-dessus de l'hibernaculum (Kapitonov 1960, Kizilov & Berendyaev 1978). De nombreuses particularités comportementales et des adaptations écophysiologiques améliorant la balance énergétique ont été déjà mention-nées. La vie à haute altitude ou latitude influence aussi l'intensité de la reproduction qui est nettement supé-rieure chez la marmotte rouge (M. caudata) de la ceinture alpine que chez celle de la zone de steppe (Kizilov & Berendyaev 1978). La gestation des femelles de la marmotte à tête noire et des autres marmottes asiatiques se passe en partie dans les terriers d'hiber-nation. La dispersion des animaux matures est retardée d'un à 3 ans dans l'extrême nord des deux continents et aux altitudes élevées, les subadultes survivent mieux dans leur famille ce qui compense le retard du début de la reproduction (Barash 1989, Bibikov 1989). Chez M. flaviventris, les populations d'altitude, présentent la polygamie et des interactions sociales complexes entre le mâle dominant et différents groupes de femelles et leurs jeunes (Armitage 1962, Barash 1989).
Activités de surface, énergétique et hibernation sont en corrélation étroite dans le cycle annuel des mar-mottes. Au cours de la période initiale d'hibernation et des derniers mois avant l'émergence des marmottes, un nombre important d'entre elles sont somnolentes. La période de torpeur profonde elle-même n'excède pas la moitié de la durée de l'hibernation. La durée du stockage de l'énergie est réduite à la seconde partie de l'été alors que les dépenses lipidiques s'étendent sur plus des 3/4 de l'année, incluant les 2-3 mois du printemps.
La durée des périodes du cycle annuel et des éta-pes de l'hibernation varient selon les espèces et les populations géographiques. La durée de l'hibernation augmente dans les conditions d'été court des hautes montagnes et de l'extrême nord. Il y a de nombreuses adaptations écologiques et comportementales : parcel-lisation de la distribution des terriers d'hivernage, construction des terriers (profondeurs, bouchons, etc.) et spécialisation de l'alimentation et de la reproduction.
*****
| Marmot age | Cellar const. T -5deg.C | Cool chamber -2 +4deg.C | Warm chamber +5 +11deg.C | ||||||
| I | II | III | I | II | III | I | II | III | |
| young | 14 | 0-2.3 | 0-2 | 6 | 0.2-10 | 0.2-5.7 | 10 | 0.6-9 | 0.6-10.5 |
| 1 | 0.9 | 3.6 | 2.2 | 2.9 | 3.3 | ||||
| adult and | 38 | 1.2-14.6 | 1.1-5 | 20 | 0.1-10.9 | 0.1-4 | 17 | 1.9-24.3 | 0.4-8 |
| subadult | 4.2 | 1.8 | 4.8 | 1.6 | 11.4 | 3.8 | |||
(I) Nombre d'animaux /Number of animals; (II) Perte journalière moyenne de masse en g / Average of daily mass loss, g; (III) Perte moyenne par décade de masse (% du poids initial en début d'hibernation) / Average of decade mass loss (per cent of the initial body mass at the onset of hibernation). Numérateur / Numerator: limites / limits; dénominateur / denominator: moyenne / mean.
1. M. marmota; 2. M. bobac (plain subspecies); 3. M. bobac centralis; 4. M. b. sibirica; 5. M. himalyana; 6. M. caudata; 7. M. menzbieri; 8. M. camtschatica (all subspecies); 8a. M. c. camtschatica; 8b. M. c. bungei; 8c. M. c. doppelmayeri; 9. M. flaviventris; 10. M. monax; 11. M. broweri; 12. M. caligata (+ M. olympus + M. vancouverensis).
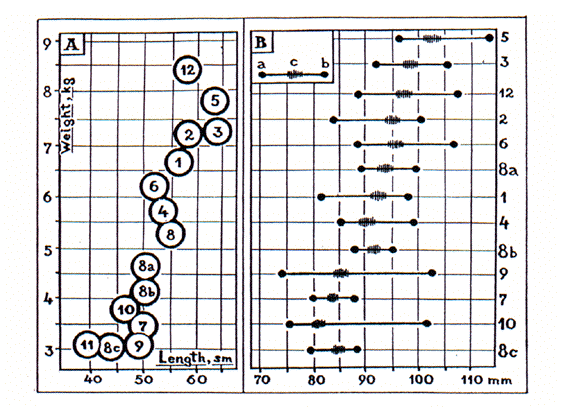
Figure 2. Cycle annuel de la marmotte du Tien Shan dans la zone des prairies-steppes du Tien Shan.
Annual cycle of the Tien-shanian marmot in the meadow-steppe zone of Tien Shan.
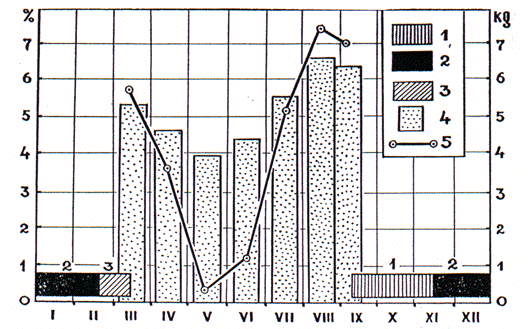
La plupart des animaux sont : 1. somnolents en automne ; 2. torpides ; 3. somnolents à la fin de l'hiver ; 4. masse moyenne des mâles adultes ; 5. moyenne de la graisse périviscérale (% de la masse corporelle).
Most animals are: 1. drowsy in autumn; 2. torpid; 3. drowsy in late winter; 4. average weight of the adult males; 5. average index of perivisceral fat (% of the body weight).
Marmots (Marmota), widely distributed in the Holarctic region; inhabit almost all natural zones from the Arctic Sea coast to the high mountains of Central Asia. Due to their extensive range marmots may seem to be animals of great plasticity in foraging, activity and choice of its habitat in particular. In reality, these colonial hibernating rodents are specialised herbivores with the typical behaviour of the inhabitants of open landscapes. They need not only a good view of the surroundings, but also a fairly thick layer of fine soil for deep wintering burrows.
The periods of surface activity and hibernation considerably vary among the members of the genus. Hibernation period in the American M. monax is very short (2-3 months) or even almost absent, whereas in M. c. camtschatica it lasts up to 8-9 months. Such considerable differences in the annual cycle are ex-plained by their living conditions. Dependence on the intensity of heat loss and of body size of the animal was found long ago. It is relatively less in larger animals (Morrison 1960, cit. Kalabukhov 1985). Panteleev (1983) noted that considerable heat loss results in selection of forms with high intensity metabolism. From this point of view increase in body size and weight of the marmots is supposed to occur as an adaptation to severe living conditions.
For the first time we make an attempt to compare body size of different marmot species to their annual cycle and adaptations to particular living conditions, and to judge connections between energetic and animal size. In North America the most part of the range is inhabited by the relatively more ancient marmots of the Marmota group (Gromov et al. 1965, Hoffman 1968). The recent forms of the bobac group that appeared in the late Pleistocene and Holocene prevail in Eurasia (Gromov et al. 1965, Bibikov 1967). More ancient forms inhabit the Alps and Carpathians (M. marmota) and Central Asia (M. caudata and others).
In our study we applied to the body size as a more often used morphological characteristic. The comparison between marmots according to their body length and weight (Fig. 1A) is performed for the first time as for the condilobasal length of the skull (Fig. 1B) on which the errors of the measuring procedure seem to be minimal. Not only measurements of the classic keys (Howell 1915, Ognev 1947, Gromov et al. 1965, Lee & Funderburg 1982, Nowak 1991), but also the measurements of particular Eurasian marmot species taken by zoologists (Petrov 1960, Tkachenko 1962) are used in the Fig. 1. Naturally, measurement techniques and sampling are not identical in different studies; I also showed certain personal preference while selecting the data of particular authors.
Mean values for each of the considered species were calculated out of not less than 10 measurements. In some cases the data of different authors were not the same, so there are some problems with their reliability, particularly as regards body length and weight of the animals. The small marmots (M. menzbieri, M. flaviventris, M. broweri and M. camtschatica, mostly its small Yakut and particularly Barguzin subspecies) as well as the large species (M. himalayana, M. caligata and M. bobac) inhabit both Asia and America (Fig. 1A). The comparison of the condilobasal length of the skull (Fig. 1B) also confirms the existence of marmots of different sizes and the gradual transition from small animals to large ones on both continents.
We pay a special attention to the presence of geographical and population differences in the size of some marmots considered as a single species. They may be one-and-half fold, for example, in the yellow-bellied marmot (M. flaviventris) from the various parts of its range and different altitudes and even two-fold in the black-capped marmot subspecies (M. camtshatica). The animals from different populations, American M. flaviventris for example, can be 1,5 times shorter and even more than 1.5 times lighter than the marmots of "caligata" group. The same phenomenon is observed in the pair of Palearctic species, the Menzbier's marmot and the bobac.
As far as geography and ecology are concerned, no attempts to compare the marmots from Asia and America are known to me. So far, western zoologists are almost unaware of the biology of Eurasian species. Our specialists almost have not discussed the problem either. This is the possible reason for the absence of any discussion on the origin of the wide overlapping of the species ranges in America and their allopatry in Europe and Asia. There are also some more differences in the biology of marmots, but the similarity in certain details, e.g., foraging, activity, population structure or behaviour are more evident. It is remarkable, that the scope of the morphological and ecological charac-teristics of the animals belonging to the same species but inhabiting various parts of the range or different altitudes is sometimes greater than that of the different European or the North American marmot species.
The structure of the annual cycle is illustrated by that of the Tien-Shan marmot (M. bobac centralis) inhabiting the mountain meadow-steppe zone. The accumulation of new fat reserves, illustrated by the increase of the index of the perivisceral fat (in % of body weight) begins only in June and gets most intensive in July and August (Fig. 2, curve). Three subsequent stages of the hibernation period can be distinguished, which differ in hibernaculum conditions, behaviour and physiology of the animals.
In the plugged hibernacula marmots find themselves in full darkness, under low temperature and high humidity and carbon dioxide level and shortage of oxygen... The increase of the carbon dioxide content in a hibernaculum, according to the self-narcosis theory by Dubois (1896) promotes the onset of torpor (Bibikov 1967, p.72).
It is interesting to notice that the deep torpor in the captive gray marmots and tarbagans began only in late October or even in December, i.e. much later than in field conditions. The recent experiments confirm the gradual transition (during October and November) to the profound torpor. The black-capped marmots became torpid in captivity 1.5-2 months later than in nature. In November their body temperature dropped, the duration of torpor bouts increased while the intervals between them became shorter (Vasilyev 1989). The experiments with M. bobac also show some changes (Rymalov & Maximov, 1992).
The
2nd stage.
The 2nd stage of the deep torpor lasts for 2 to 3 months, from late
November till late February (Fig. 2.2). Most of the animals are torpid and the
number of the dozing marmots is very low. Recent 7-year study of the
temperature regime and the oxygen content in six wintering burrows (Arnold et
al. 1991) has shown that 2.5 months before the onset of profound torpor the
temperature in the hibernacula at a depth of 1-3 m abruptly dropped (from
12deg. to 5deg.C) and thereafter this process became gradual (approximately
1deg. per month).
The determination of regular breaks of the torpor occurring all along
hibernation, a very impor-tant phenomenon, was discovered for the first time in
the golden-mantled ground squirrel (Citellus lateralis; Pengelley &
Fisher 1961). An assumption about the presence of the breaks in the marmot
hibernation was put forward by Berendyaeva et al. (1967) who placed
telethermometer AM-2M in hibernaculum of the Tien-Shanian marmot located 3400 m
above s.l. Thermometer reading taken three times a day showed temperature
fluctuations. According to the authors' opinion, temperature rises depend on
the arousal of the hibernating marmots every 10 days. After December-January,
the torpor periods of the black-capped marmots became more prolonged: they
lasted for 15-16.5 days in the adult female, 17 days in the yearlings and
20.5-22 days in the infants (Vasilyev 1989).
The 3rd stage. This is the period of arousal, mating and winter burrow leaving. This stage is less prolonged (March or April-May) in the case of short summer (Fig. 2,3). It is characterised by more frequent breaks in torpor bouts and increased number of drowsy animals. It remains unknown how the ventilation of the chamber is performed in the constant presence of at least one or two active or drowsy reproductive marmots. The problem of energy expenditure during the annual cycle is to be discussed in more detail. The amount of subcutaneous and perivisceral fat has been shown to vary in the studied Eurasian marmots from 20 to 30 % of the body weight on the eve of hibernation. Only in the black-capped marmots from Yakutia the value of this index appeared to be about 35%, which is slightly higher than in all other investigated marmots. Not only the amount of fat, but also the differences in its composition are of great importance. Data on the acid composition of marmot fat are incomplete, though it is known that there is more linoleic acid in the fat of the susliks inhabiting mountains than of those from the lowlands (Kalabukhov, 1985). The physicochemical constants of fat are also variable (Bliznetzov 1966).
Little is known about the dynamics of the energy expenditure over the hibernation stages. In M. monax the total weight loss due to the energy expenditure was 1245 g; 41.6% of weight being lost in the wintering burrow before the onset of hibernation, 16.9% - during hibernation itself and 41.6% upon arousal, but before the beginning of the active foraging in March (Bailey & Davis 1965). During the active period the Eurasian marmots that are totally devoid of food lose 3.76% of their body weight per day. These indices are between 0.24 and 0.35% for the drowsy rodents and only between 0.02 and 0.12% for torpid individuals. In our experiments the Tien-Shan marmots lost 0.09 to 0.18% of their original body weight per day on average, which corresponds to an absolute mass loss of 1.0 to 4.2 g (Bibikov 1967).
The experiments on the influence of hibernating conditions on energy expenditure in the Tien-Shan marmots showed that the weight loss was greatest under fluctuating and relatively high temperature (8+/-3deg.C). The animals saved more fat while hibernating under a constant temperature (5deg.C) in the cellar (Table 1).
In our experiments we paid attention to the adap-tability of continuous hibernation and more economic energy expenditure by the young animals. We supposed that the fatter adults wake up more frequently, warming the hibernaculum and hence promoting successful wintering of the infants. Our assumptions have recently been supported by the series of studies carried out in Bavaria (Arnold 1990, Arnold et al. 1991).
However, the possibility of survival under conditions of short summer is promoted by the presence of numerous adaptations. The first is the choice of territory in suitable biotopes with food in spring and thin layer of fine soil (Bibikov & Stogov 1957, Barash 1973, 1989). Under a shortened vegetation period the specialisation in juicy food (roots and bulbs) becomes more pronounced, e.g., in the Menzbier's, black-capped and Olympic marmots. On the contrary, the Kazakhstan bobacs inhabiting dry steppe with frequent droughts adapted to the consumption of rough food, such as dry grass and even the twigs of shrubs.
A set of adaptations to the extreme conditions is connected with the complication of the wintering burrows and the better isolation of hibernaculum: the favourable location of the burrow in the mezorelief, a great depth, a thick bedding, numerous ground "plugs" (there are also additional grass "plugs" in the burrows of the Yakut marmots) and even a "mound" above hiber-naculum (Kapitonov 1960, Kizilov & Berendyaev 1978). Numerous behavioural peculiarities and ecophysiolo-gical adaptations aimed at the improvement of the ener-gy balance have been already mentioned. Living in high altitudes and latitudes also influences the intensity of the reproduction (e.g., it is considerably higher in the red marmot from the alpine belt than in the animals of the same species from the steppe zone (Kizilov & Berendyaev 1978). The pregnancy of the female black-capped and other Asian marmots partly takes place in wintering burrows. The dispersal of the mature animals delayed for 1-3 years in the Far North of both continents and in high altitudes, the subadult animals survive more successfully in the parental family which compensates for the late beginning of the reproduction (Barash 1989, Bibikov 1989 and others). In high moun-tain populations of yellow-bellied marmots polygamy and rather complicated social interactions between the dominant male and several groups consisting of females and young are observed (Armitage 1962, Barash 1989).
The surface activity, energetic and hibernation are closely connected in the annual cycle of the marmots. In the initial period of hibernation and during the last month before marmots' emergence on the surface a lot of animals remain drowsy. The period of profound torpor itself (the 2 nd stage of hibernation) does not exceed one half of the hibernation period. The time of energy storage are restricted to the second half of the summer, while fat expenditure lasts for more than 3/4 of the year, including 2-3 spring months.
The proportion between the duration of periods of the annual cycle and the stages of hibernation varies in different species and in geographical populations. The duration of total hibernation period and its stages increases under the conditions of short summer in the high mountains and in the Far North. There are numerous ecological and behavioural adaptations: the patchiness of the distribution in wintering burrows, the construction of the burrows themselves (the depth, the plugs, etc.) and specialisation in foraging and in reproduction.
ARMITAGE K.B. 1962. Social behavior of a colony of the yellow-bellied
marmot (Marmota flaviventris). Anim. Behav., 10: 319-331.
ARNOLD W. 1990. The evolution of marmots sociality: I, II. Behav.
Ecol. Sociobiol., 27: 229-246.
ARNOLD W., HELDMAIER G. & ORTMANN S. 1991. Ambient temperatures in
hibernacula and their energetic consequences for Alpine marmots (M.
marmota). J. Term. biol., 16: 223-226.
BAILEY E.D. & DAVIS D.E. 1965. The utilization of body fat during
hibernation in woodchucks. Can. Journ. Zool., 43: 701-707.
BARASH D.P. 1973. The social biology of the Olympic marmot. Anim. Behav.
Monogr., 6: 171-249.
BARASH D.P. 1976. Pre-hibernation behavior of free living hoary marmots, M.
caligata. J. Mamm., 57: 172-185.
BARASH D.P. 1989. Marmots. Social behavior and ecology. Stanford Univ.
Press, Stanford, 360 pp.
BERENDYAEVA E.L., KULKOVA N.A. & MIKHAILUTA A.A. 1967. On the temperature
regime in the nest chambers of the gray marmots in Tien Shan. Proc. 5
scientific conf. antiplague establ. of Middle Asia and Kazakhstan,
Alma-Ata: 97-99 (in Russian).
BIBIKOV D.I. 1967. Mountain marmots of Central Asia and Kazakhstan.
Moscow, Nauka, 198 pp. (in Russian)
BIBIKOV D.I. 1989. The marmots. Moscow, Agropromisdat, 255 pp. (in
Russian)
BIBIKOV D.I. & STOGOV I.I. 1957. Ecological mechanisms of the statial
distribution of the gray marmots (M. b. baibacina) in Tien Shan.
Byul. Moskovskogo Obschestva Ispytatelei Prirody, otdel biologii, 62:
13-18. (in Russian)
BLIZNETZOV I.Ya. 1966. Some eco-physiological peculiarities of the torpid
gray marmots and yellow susliks. Ref. cand. diss. Frunze, 19 pp. (in
Russian)
DUBOIS R. 1896. Physiologie comparée de la marmotte. Etude sur le
mécanisme de la thermogénese et du sommeil chez les
mammifères. Annales de L'Universite de Lyon, 25.
GROMOV I.M., BIBIKOV D.I., KALABUHOV N.I. & MEIER M.N. 1965. Ground
squirrels (Marmotinae). In USSR fauna. Mammals, V.3, 2. New ser.,
N 68, M.-L., Nauka, 468 pp. (in Russian)
HOFFMANN R.S. 1968. Russian Science: a personal View. World Book Encyclopedia,
Science Year: 228-239.
HOWELL A.H., 1915. Revision of the American marmots. U.S. Government
printing office, Washington, D.C.
KALABUKHOV N.I. 1985. Mammal hibernation. Moscow, Nauka, 260 pp. (in
Russian)
KAPITONOV V.I. 1960. Biological sketch of the black-capped marmot (M.
camtschatica Pall.). Zool. zh., 36: 448-457. (in Russian)
KAPITONOV V.I. 1978. The black-capped marmot. The Menzbier's marmot. In The
marmots. Distribution and ecology, R.P.Zimina ed., Moscow, Nauka: 126-157,
178-209. (in Russian)
KIZILOV V.A. & BERENDYAEV S.A. 1978. The red marmot. In The marmots.
Distribution and Ecology, R.P. Zimina ed., Moscow, Nauka: 79-116. (in
Russian)
LEE D.S. & J.B. FUNDERBUR g, 1982. Marmots. In:Wild Mammals of North
America, T. Chapman & G. Feldhamer eds, Hopkins Univ. Press, Baltimore:
176-191.
NOWAK R.M., 1991. Marmots. In Mammals of the world, V.1., Hopkins Univ.
Press, Baltimore & London: 567-569.
OGNEV S.I. 1947. Mammals of the USSR and neighboring countries. Rodents,
5. Moscow, Akad. Nauk USSR, 800 pp. (in Russian)
PANTELEYEV P.A. 1983. Bioenergetics of small mammals. Moscow, Nauka, 269
pp. (in Russian)
PENGELLEY E.T. & K.C. FISHER, 1961. Rhythmical arousal from hibernation in
the golden-mantled ground squirrel, Citellus lateralis. Can. J.
Zool., 39: 105-120.
PETROV B.M. 1960. On ecology of Menzbier's marmot. Izv. Biol., Tashkent,
3: 42-47. (in Russian)
RYABOV N.I. 1948. Materials to biology of Transbaicalian marmot in winter
period. Zool. zh., 27(3): 245-256. (in Russian)
RYMALOV I.V. & MAXIMOV A.Yu. 1992. Hibernation of marmots in captivity.
Proc. 1st Intern. Marmots Symp. on Alpine Marmot, Bassano B.,
Durio P., Gallo Orsi U., Macchi E. eds., Torino Univ., Italy: 63-66.
TKACHENKO A.A. 1962. Biology and agricultural significance of the steppe
marmot in the virgin lands of Kazakhstan. Ref. cand. diss., Moscow, Moscow
Univ., 22 pp. (in Russian)
VASILYEV V.N. 1989. Ecology of hibernation in black-capped marmot (M.
camtschatica). Ref. cand. diss., Akad. Nauk SSSR, Moscow, 22 pp. (in
Russian)
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |