
Trente-six espèces de Siphonaptères ont été
trouvées sur les marmottes grises (Marmota baibacina) et à
longue queue (M. caudata) ainsi que dans leurs nids, dans les foyers
pesteux (Tien Shan central, Alaï, monts Gissar et Talas), et hors de
ceux-ci (Pamir oriental). Huit à 21 espèces de puces existent
dans chaque région, mais seules Oropsylla silantiewi,
Citellophilus lebedewi et Rhadinopsylla li ventricosa, sont
présentes partout. L'abondance des parasites dépend de
l'altitude. Pulex irritans est un élément important de
cette faune dans certains foyers. Les 32 espèces restantes
représentent 0,3% des puces capturées. De tels résultats
montrent un contact parasitaire faible entre les marmottes et d'autres animaux
partageant les mêmes milieux.
Mots clés : Puces, Siphonaptères, marmottes, foyer de
peste, Tien Shan, Pamir, Alaï, Gissar, Talas.
Thirty-six species of Siphonaptera were found as a whole on grey
(Marmota baibacina) and long-tailed (M. caudata) marmots
as well as in their nests in the plague foci of Central Tien Shan, Alai, Gissar
and Talas mountain ridges as well as in Eastern Pamir where no plague was
discovered. From 8 to 21 species occurred in each separate region, but only
Oropsylla silantiewi, Citellophilus lebedewi and Rhadinopsylla li
ventricosa, specific parasites of marmots, were met everywhere. Number of
most parasites depends on mountain altitudinal zones. Pulex irritans is
also an important component of the marmot flea fauna in some foci. The rest of
32 species comprised 0.3 per cent of all collected fleas. Such findings prove a
weak parasitic contact between marmots and other animals living in the same
habitats.
Key-words: Fleas, Siphonaptera, Marmots, plague foci, Tien
Shan, Pamir, Alai, Gissar, Talas.
| Ics = | 2a ---------------- (a+b) + (a+c) | X 100 |
où a est le nombre d'espèces communes aux deux faunes comparées, b, le nombre d'espèces de la faune B et c, celui de l'espèce C.
D'un autre côté, les puces spécifiques des mar-mottes ont été trouvées sur 9 espèces de rongeurs et 5 de carnivores. Parmi les 23.722 puces récoltées, celles des rongeurs de ces régions ont formé 0,3% du total. Mais sur les 1.967 exemplaires provenant des carnivores, la part des puces de marmottes a été plus importante et a atteint 17,1%. Selon certains auteurs (Petrov, Schmuter 1958), ceci peut s'expliquer par l'écologie de ces hôtes. Lorsque les prédateurs, surtout les mustélidés, visitent les nombreux terriers de marmottes, ils subissent l'attaque des puces et les collectionnent sur de vastes territoires. Il doit être noté que la proportion de P. irritans, fréquemment associée aux grands carnivores (Lewis 1972), a été plus faible que celle des puces spécifiques des marmottes et a atteint 10% seulement.
Il découle de ces faits que O. silantiewi, C. lebedewi et R. li ventricosa montre une spécificité relativement forte pour leurs hôtes. Ceci est confirmé par de nombreuses observations de terrains qui indi-quent que les puces de marmottes n'abandonnent leur hôte tué ou mort qu'après un long laps de temps (Bibikov et al. 1973).
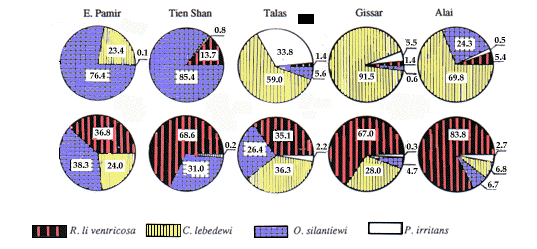
Les proportions des espèces dominantes de ces faunes sur les marmottes ne sont pas constantes et dé-pendent de l'altitude. O. silantiewi prédomine sur les marmottes (Fig. 2) des ceintures alpines froides (3.000-4.000 m au-dessus du sol) du Tien Shan et du Pamir (76-85%), alors que C. lebedewi est dominante (59-91%) dans les ceintures plus chaudes de prairie-forêt-steppe (2.200-3.000 m) des sommets de l'Alai, du Gissar et du Talas.
Les steppes montagneuses sèches (jusqu'à 2.200 m) et les parties inférieures des ceintures citées précé-demment sont les aires préférées par P. irritans qui atteint jusqu'à 30% dans certaines zones (Yakunin et al. 1985). R. li ventricosa n'est pas un parasite fréquent dans la fourrure des marmottes, mais il prédomine (Fig. 2) dans leur terrier (jusqu'à 83,8%).
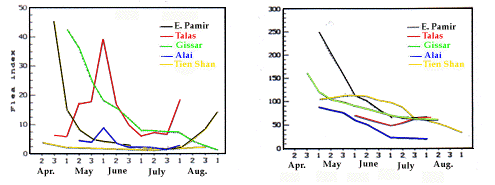
Le nombre total de puces dans une famille de marmottes est de l'ordre de quelques dixièmes de celui des insectes qui sont ses hôtes et de quelques centièmes de ceux occupant la litière d'un terrier hivernal ou de 2-10 terriers estivaux et n'excède pas 800. Ce nombre sur l'animal ou dans le terrier varie durant la saison d'ac-tivité des marmottes. Les données concernant la période d'hibernation ne sont pas connues. Le nombre moyen de puces par hôte ou puce index (Fig. 3) est maximum au printemps et au début de l'été et minimum au milieu de l'été et en automne (Fig 4). En général, cependant, ce nombre par marmotte est plusieurs fois inférieur à celui des mois comparables dans les nids. Les variations con-trastées de ce nombre de puces de la fourrure sont dues à O. silantiewi et C. lebedewi dont les adultes atteignent un pic d'émergence au cours du printemps. Le nombre élevé de puces au printemps dans les nids est causé par R. li ventricosa qui éclosent à cette saison aussi.
Il ne peut y avoir aucun doute sur l'importance d'O. silantiewi, C. lebedewi, R. li ventricosa dans la pro-pagation de la maladie. Selon Dmitrievskaja et al. (1961), le nombre de germes de peste dans les cultures, isolées à partir de chaque espèce de puce, serait dépen-dant du nombre de puces dans la nature et de sa propor-tion dans la faune à un moment donné. Au début de l'été, O, silantiewi est d'un point de vue numérique et épizootique la plus importante des trois espèces dans la fourrure des marmottes, alors que R. li ventricosa est dominante parmi celles infestant les nids de marmottes.
*****
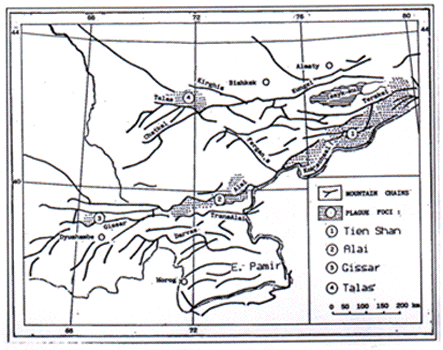
| Mountain region | Enzootic area sq. km | Date of discovery | Elevation above sea level in m | Marmot species |
| C. Tien Shan | 20,000 | 1914 | 2,600-4,000 | grey (M. baibacina) |
| Alaï | 2,800 | 1948 | 2,200-3,800 | long-tailed (M. caudata) |
| Gissar | 300 | 1970 | 2,250-3,400 | long-tailed |
| Talas | 2,500 | 1977 | 2,250-3,200 | long-tailed |
| E. Pamir | 3,600-5,000 | long-tailed |
| Area | Host nb. | Total number of mamot fleas | Nest nb. | Total number of nest fleas | ||||||||
| O. silantiew | C. lebedewi | R. ventricosa | P. irritans | Others | O. silantiew | C. lebedewi | R. ventricosa | P. irritans | Others | |||
| Tien Shan | 58,168 | 51,460 | 502 | 8,235 | 34 | 410 | 9,321 | 7 | 20,613 | 112 | ||
| Alai | 7,772 | 3,305 | 9,487 | 732 1 | 65 | 38 | 81 | 82 | 1,007 | 1 | 30 | |
| Gissar | 3,308 | 299 | 444,014 | 667 | 2,548 | 477 | 41 | 174 | 1,024 | 2,450 | 5 | 5 |
| Talas | 3,160 | 2,206 | 23,296 | 671 | 13,290 | 39 | 41 | 572 | 787 | 760 | 1 | 47 |
| Pamir | 17,479 | 87,811 | 26,948 | 116 | 16 | 85 | 3,250 | 2,032 | 3,118 | 75 | ||
| Espèces / Species | 1 | 2 | 3 | 4 | 5 |
| Pulicidae | |||||
| 1. Ctenocephalides canis (Curtis 1826) | + | ||||
| 2. Pulex irritans Linné 1758 | + | + | + | + | |
| Vermipsyllidae | |||||
| 3. Chaetopsylla homoea homoea Rothshilds 1906 | + | + | + | ||
| Hystrichopsyllidae | |||||
| 4. Neopsylla mana Wagner 1927 | + | + | |||
| 5. N. meridiana Tiflov & Kolpakova 1937 | + | + | + | + | |
| 6. N. pleskei ariana Ioff 1946 | + | + | + | ||
| 7. N. teratura Rothshild 1913 | + | + | |||
| 8. Paraneopsylla ioffi ioffi Tiflov 1937 | + | + | + | + | |
| 9. Rhadinopsylla angusta Tiflov 1937 | + | ||||
| 10. R. dahurica tjanshan Ioff & Tiflov 1946 | + | ||||
| 11. R. li murium Ioff & Tiflov 1946 | + | ||||
| 12. R. li ventricosa Ioff & Tiflov 1946 | + | + | + | + | + |
| 13. Stenoponia ivanovi Ioff & Tiflov 1934 | + | ||||
| 14. S. suknevi Ioff & Tiflov 1934 | + | ||||
| Leptopsyllidae | |||||
| 15. Amphipsylla anceps Wagner 1930 | + | + | + | ||
| 16. A. asiatica (Ioff 1928) | + | + | |||
| 17. A. montana Argyropulo 1946 | + | ||||
| 18. A. phaiomydis phaiomydis Ioff 1946 | + | + | |||
| 19. A. primaris primaris J. & R. 1915 | + | + | + | ||
| 20. Frontopsylla ambigua Fedina 1946 | + | ||||
| 21. F. elata glabra Ioff 1946 | + | + | + | + | |
| 22. F. elata neutra Mikulin 1947 | + | ||||
| F. frontalis alatau Fedina 1946 | + | + | + | + | |
| 24. F. ornata Tiflov 1937 | + | ||||
| 25. F. protera Wagner 1933 | + | ||||
| 26. F. tjanshanica Schwarz 1953 | + | ||||
| 27. Leptopsylla nana Argyropulo 1946 | + | ||||
| 28. L. nemorosa (Tiflov 1937) | + | ||||
| Ceratophyllidae | |||||
| 29. Callopsylla capsia (Ioff & Argyropulo 1934) | + | + | + | + | + |
| 30. Ceratophyllus avicitelli Ioff 1946 | + | + | + | + | |
| 31. Citellophilus lebedewi princeps (Ioff 1946) | + | + | + | + | + |
| 32. C. relicticola (Fedina 1946) | + | ||||
| 33. Malaraeus penicelliger syrt (Ioff 1946) | + | ||||
| 34. Monopsyllus sciurorum asiaticus (Ioff 1936) | + | ||||
| 35. Oropsylla ilovaiskii Wagner & Ioff 1926 | + | ||||
| 36. O. silantiewi (Wagner 1898) | + | + | + | + | + |
Table 4. Coefficients of similarity between local marmot flea faunas (Czecanowski & Sørensen's method)
| Region | Species Nb. in local fauna | Tien Shan | Alai | Gissar | Talas | E. Pamir |
| Tien Shan | 21 | 12** | 10 | 14 | 7 | |
| Alai | 15 | 66.7* | 10 | 5 | ||
| Gissar | 18 | 51.3 | 60.6 | 10 | 5 | |
| Talas | 20 | 68.3 | 82.8 | 52.6 | 7 | |
| Pamir | 8 | 48.3 | 52.2 | 47.6 | 50.0 |
Figure 1. Plague enzootic areas in the mountains of Central Asia
|
Seasonal change of total flea indices on marmots
Figure 4. Variation saisonnière du nombre total de puces dans les
nids de marmottes | |
| Ics = | 2a ---------------- (a+b) + (a+c) | X 100 |
where a: number of species common to both faunas compared, b: number of species belonging to B fauna solely, c: number of species belonging to C fauna solely.
Czecanowski and Sørensen's coefficient of simil-arity (Tab. 4) is highest between Alai and Talas faunas (82.8% of species common to both faunas compared) and is the lowest between Pamir and Gissar (47.6 % of common species). Only O. silantiewi, C. lebedewi, R. li ventricosa and C. caspia, were found in all compared regions. The first three are specific and predominant marmot parasites. They respectively represented 49.3%, 33.6% and 11.9% of the total. Pulex irritans, the so-called human flea, is also an important component of the marmot flea fauna in some foci, and represented 4.9% of the total. The rest of 32 species, including C. caspia, comprised only 0.3% of all collected fleas. All of them are chance parasites of marmots; and such fin-dings show weak parasitic contacts of marmots with suslics, variety of small murine and microtine rodents, feral carnivores and hole dwelling birds.
On the other hand, the specific marmot fleas were found on 9 species of rodents and 5 species of car-nivores. Of 23,722 fleas collected from rodents in these regions they comprised 0.3% of the total. But among 1,967 specimens combing from carnivores the share of marmot fleas was much higher, and it comprised 17.1%. According to some authors (Petrov & Schmuter 1958) this fact can be accounted for ecology of these hosts. While visiting numerous marmot burrows, the pre-dators, mainly musteline species, undergo fleas attacks and collect these insects from vast territories. It should be noted that the proportion of P. irritans, frequently associated with large carnivores (Lewis 1972), was smaller than those of specific marmot fleas and comprised 10% only. It follows from these facts that O. silantiewi, C. lebedewi and R. li ventricosa show relatively high host specificity. This ability is supported also by many field observations which show that marmot fleas don't leave their shot or died hosts over a long period of time (Bibikov et al. 1973).
The proportions of prevailing species in the mar-mot flea faunas are not constant, and they depend on altitudinal zonality. O. silantiewi predominates on mar-mots (Fig. 2) in the cold alpine belts (3,000-4,000 m above sea level) of Tien Shan and Pamir (76-85%), while C. lebedewi predominates (59-91%) in the rela-tively warmer meadow-forest-steppe belts (2,200-3,000 m) of Alai, Gissar and Talas ridges. Dry mountain steppes (up to 2,200 m) and the lower part of the above belts are preferred areas for P. irritans that comprises up to 30% in some localities (Yakunin et al. 1985). R. li ventricosa is not a dominant parasite in marmot fur, but it predominates (Fig. 2) in their nests (up to 83.8%).
The total number of fleas within a marmot family range consists of a few tens of insects which are on the hosts and a few hundreds of insects which are in the nest linings of one hibernal or in 2-10 estival burrows and does not exceed 800. This number on the host body and in its nests fluctuates during the season of ground activity of marmots. Data concerning over-wintering period are absent. The average flea number per host or so-called flea index (Fig. 3) is the greatest in spring and early summer and is the lowest in mid-summer and fall. Flea indices in the nests reflect to a certain extent the flea indices on the hosts (Fig. 4). In general, however, fleas per marmot were several-fold fewer than fleas per nest for the comparable months. The sharp monthly fluctuations in number of hair fleas were due to O. silantiewi and C. lebedewi that reach a peak of adult emergence during spring. The higher spring numbers in nests are caused by R. li ventricosa that hatches in this season also.
There can be no doubt about the importance of the biology of O. silantiewi, C. lebedewi, R. li ventricosa in the spread of the disease. The number of plague cultures isolated from separate flea species is thought to be dependant largely upon their numbers in nature and upon their percentages in the flea fauna at a given time. (Dmitrievskaja et al. 1961) In the early summer O. silantiewi is numerically and epizootically the more important of the three above species combed from marmots, while R. li ventricosa is predominant among infected nest fleas collected in the season.
SøRENSEN Y. 1948. A new method of equal amplitude in plant sociology
based on similarity of a species content and its application to analysis of the
vegetation on Danis commons. Kgl. Dan. vedenskab. selskab biol skr., 5:
1-34.
YAKUNIN B.M., A.V. SCHWARZ & J.S. SLJUNKIN 1985. On the flea fauna of Talas
plague focus. In Urgent problems of surveillance in the natural plague
foci, Stravopol, 121-122. (In Russian)
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |