![]()
Le partage des ressources, mesuré par le chevauchement des espaces
utilisée, est orienté principalement envers la parenté
proche et ne soutient pas l'hypothèse de la distribution proportionnelle
des bénéfices selon laquelle ceux-ci sont distribués en
proportion du degré d'apparentement. La formation matrilinéaire
est le mécanisme social par lequel les femelles de la marmotte à
ventre jaune tentent de maximiser la valeur sélective globale en
exagérant la composante directe de la valeur sélective.
Mots-clés : Marmotte à ventre jaune, valeur sélective globale, utilisation
de l'espace, apparentement, valeur sélective directe,
matrilinéaire.
Resource sharing, measured as space-use overlap, is directed primarily
toward close kin and does not support the proportional distribution of benefits
hypothesis in which benefits are distributed in proportion to the degree of
relatedness. Matrilineal formation is the social mechanism whereby female
yellow-bellied marmots attempt to maximize inclusive fitness by emphasizing the
direct fitness component.
Key words:Yellow-bellied marmot, inclusive fitness, space-use, relatedness, direct fitness,
matriline.

Les marmottes sont des animaux intéressants pour explorer les rapports entre apparentement et valeur sélective globale d'une part, du fait de la variété de leurs systèmes sociaux (Armitage 1992) et d'autre part, parce que les liens de parenté de tous les membres d'un groupe social peuvent être déterminés et les relations sociales de chaque animal observé directement.
Ce travail rend compte des relations entre le partage des ressources et la parenté d'une colonie de marmottes à ventre jaune (Marmota flaviventris) chez qui un nombre important de degrés d'apparentement apparaît.
Le rapport prévisible entre les liens de parenté et le recouvrement de l'espace utilisé a été calculé sur la base de la valeur moyenne de recouvrement lorsque l'apparentement r = 0,5 et en postulant que la valeur moyenne de recouvrement correspondant aux autres valeurs de r est réduite proportionnellement à celle de r. Par exemple, nous pouvons prédire que la moyenne de recouvrement pour r = 0,25 correspond à la moitié de la valeur moyenne de recouvrement lorsque r = 0,5.
Cependant, la forte variation des valeurs de recouvrement de l'espace utilisé quand r = 0,25 ou plus nécessite une explication. De façon nette, certains individus dont le r est égal à 0,3125 ou 0,25 partagent l'espace à un degré plus fort que certain individus reliés entre eux par 0,5. Pour expliquer cette forte variation des valeurs de recouvrement il est donc nécessaire d'examiner avec soin la structure et la dynamique sociale d'une population. Cet examen sera présenté dans la discussion après la description des patrons d'utilisation de l'espace par différents groupes.
Les animaux liés entre eux par 0,5 se trouvaient dans trois catégories ; mère-descendants, membres d'une même portée, et membres de portées différentes. Pour chacun d'eux, la moyenne de recouvrement diminue au fur et à mesure que les individus vieillissent (Fig. 2). Cependant, en moyenne, les membres d'une même portée maintiennent un fort recouvrement même à l'âge adulte, comme la mère et sa descendance, alors que les adultes de portées différentes, bien qu'ils soient liés à 0,5, présentent la moyenne la plus faible, même si celle-ci reste supérieure à celle des animaux dont la liaison est de 0,25 ou moins (Fig 1). Dans ces trois groupes, la variation est supérieure chez les adultes. De la même façon, l'espacement entre individus de la même portée chez la marmotte chenue (M. caligata) augmente avec l'âge (Barash 1989: 215).
Les animaux liés à 0,25 se trouvaient dans trois catégories ; demi-frères paternels, grand-mère/petits enfants, et tante/nièce (Fig. 3). Le recouvrement moyen le plus élevé (9,7%) s'observe entre adultes et jeunes (relation de type tante/nièce) et cette catégorie présente aussi la plus grande variation des valeurs. Cette variation est la même que celle observée chez des animaux reliés par 0,3125 (Fig. 1) ; tous ces individus étaient de portées différentes mais avaient le même père et n'étaient apparentés que faiblement par leurs mères. Le partage de l'espace des catégories tante/nièce décroît de jeunes aux un-an jusqu'aux adultes. La moyenne pour les adultes de la catégorie tante/nièce (3,5%) n'était que légèrement plus élevée que celle des adultes non apparentés (3,2%). De la même façon, tous les groupes de demi-frères paternels avaient un faible recouvrement, particulièrement quand un adulte ou un-an était associé avec un animal plus jeune (Fig. 3). Dans ce cas aussi, la variation était plus importante parmi les adultes. Finalement, le recouvrement de l'espace utilisé est bas entre les grand-mères et leur petits enfants jeunes.
En 1983, la marmotte 855 est devenue résidente à Lower Picnic et en 1991 a recruté 6 filles repro-ductrices (Armitage 1992). Les sept femelles ont formées 3 lignées matrilinéaires (Tableau 1). Ainsi, la matrilignée 1 comprenait la matriarche et deux soeurs de portées différentes ; la seconde matrilignée, une paire de soeurs de la même portée ; et la matrilignée 3, deux soeurs de portées différentes. Tous leurs descendants ont eut le même père et tous ces descendants sont appa-rentés aux femelles adultes au travers de leurs mères ; donc r = 0,3125. Dans chaque matrilignée, les jeunes ont présenté un fort recouvrement spatial, expliquant ainsi les fortes valeurs de la distribution (Fig. 1). Cepen-dant, les jeunes de différentes matrilignées ont présenté un taux de recouvrement faible ou nul, expliquant ainsi les valeurs faibles de la distribution. Du fait de l'exis-tence d'un plus grand nombre de valeurs pour les jeunes entre matrilignées que pour ceux d'une même matri-lignée, la valeur moyenne est relativement basse et plus faible que celle prédite à partir de l'hypothèse de la distribution proportionnelle des bénéfices.
La même formation matrilinéaire explique la grande variation des valeurs de recouvrement des espaces utilisés quand l'adulte est une tante et les jeunes ses nièces (Fig. 3). Ainsi le recouvrement était élevé entre les jeunes de 478 et leur tante 497 et entre les jeunes de 497 et leur tante 478. Au contraire, le recouvrement de l'espace utilisé était faible entre les jeunes des femelles dans une matrilignée et leurs tantes des autres matrilignées. Comme il y a beaucoup plus de comparaisons de recouvrement entre les matrilignées qu'au sein de celles-ci, la moyenne de recouvrement était seulement de 9,7%.
La même matrilignée illustre la forte étendue de la variation des valeurs de recouvrement de l'espace utilisé entre les mères/descendants adultes et les adultes compagnons de portées. Ainsi, la marmotte 855 présen-tait un important recouvrement de l'espace utilisé avec ses filles de la matrilignée 1, mais de très faibles valeurs de recouvrement avec ses filles des matrilignées 2 et 3 (Tableau 1). Au contraire, la marmotte 761 avait un fort recouvrement avec sa soeur 100, d'une portée différente, mais un faible recouvrement avec sa soeur de portée 733.
Le fort recouvrement entre un adulte et ses demi-soeurs adultes paternelles se présentait à Upper Picnic. Les soeurs de la même portée ont occupé le site en 1980 et ont établi une matrilignée. Ensuite, les descendants trois générations plus tard avaient le même père, mais différentes mères (il y avait un lien de parenté distant et très bas au travers des arrières grands-mères). Comme ils étaient dans la même matrilignée, ils partageaient l'espace. La plupart des demi-frères adultes paternels vivent dans des matrilignées différentes et le recouvrement de l'espace utilisé est très bas, d'où la valeur moyenne basse (Fig. 3).
Le fort recouvrement de l'espace utilisé par des marmottes à ventre jaune étroitement apparentées est similaire au patron d'utilisation de l'espace par l'unité familiale chez la marmotte alpine (M. marmota). La famille partage l'espace mais il y a peu de chevau-chement avec les familles voisines (Perrin et al. 1993). En contraste, il y a peu de recouvrement d'espace utilisé par les femelles de M. monax (Meier 1992), chez qui, en effet, chaque individu forme sa propre matrilignée.
Deux conclusions majeures deviennent appa-rentes. D'abord, les bénéfices ne sont pas distribués proportionnellement à la parenté, mais sont concentrés sur les parents proches, particulièrement ceux liés par 0,5. Du fait que les valeurs de recouvrement des parents liés entre eux par 0,125 ou moins sont inférieures à celles des animaux non apparentés, les parents reliés par r<0,25 peuvent être considérés comme s'ils n'étaient pas apparentés. Les adultes non apparentés ont de plus fortes valeurs de recouvrement d'espace quand ils sont en conflit et un mâle adulte réalise des excursions dans le domaine vital d'un autre et initie des comportements agonistiques, d'où résulte le déplacement de la femelle subordonnée par la femelle dominante d'une partie ou de tout son domaine vital (Armitage 1992).
Deuxièmement, l'organisation matrilinéaire est le mécanisme social par lequel les marmottes femelles à ventre jaune tentent de maximiser leur valeur sélective globale. Quoique il ne soit pas toujours clair pour quelle raison un individu particulier forme une matrilignée, l'appartenance à une matrilignée est rarement le fait d'animaux liés entre eux par r<0,5. La plupart des matrilignées sont des associations mère/fille avec des groupes soeur/soeur formant environ 22% de toutes les matrilignées (Armitage 1989). La familiarité avec les membres de la matrilignée semble être la base du lien social et dans de rares circonstances inclut des individus reliés entre eux par 0,25 mais jamais <0,25. Tous les recouvrements d'espace utilisé par des individus liés entre eux par <0,25 se produisent aux frontières des domaines vitaux (Fig. 10, Table 5 in Armitage 1989). La structure et la fonction matrilinéaire indiquent fortement que les marmottes à ventre jaune tentent de maximiser leur valeur sélective directe et que les bénéfices pour la valeur sélective indirecte sont une conséquence secondaire des stratégies de la première.
*****
| | | |
| 855 | ||
| | ||
| 761 | | |
| 100 | ||
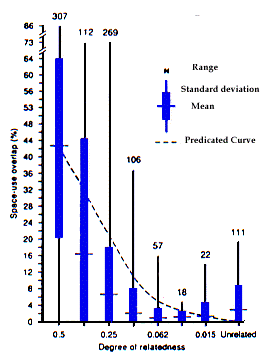
Figure 2. Recouvrement de l'espace utilisé par les marmottes à
ventre jaune apparentées à 0,5.
Space-use overlap for yellow-bellied marmots belonging to kinship groups
whose members are related by 0.5.
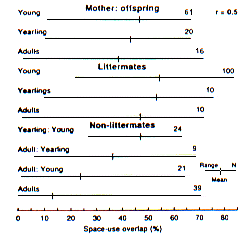
Figure 3. Recouvrement de l'espace utilisé par les marmottes à
ventre jaune apparentées à 0,25
Space-use overlap for yellow-bellied marmots belonging to kinship groups
whose members are related by 0.25.

Marmots are excellent animals for exploring the relationship between relatedness and inclusive fitness because of the variety of social systems (Armitage 1992) and because relatedness of all members of a social group can be determined and the social relations of each animal can be directly observed.
Herein I report the relationship between resource sharing and "kinship" for a colony of yellow-bellied marmots (Marmota flaviventris) in which a wide range of degrees of relatedness occurred.
A predicted relationship between relatedness and space-use overlap was calculated by using the mean value of space-use overlap for r = 0.5 as the base value and assuming that the mean value of space-use overlap for all other r values was reduced by the same proportion the r value was reduced. For example, the mean space-use overlap for r = 0.25 was predicted to be one-half the mean value for r = 0.5.
However, the wide range of space-use overlap values when r =0.25 or greater requires explanation. Clearly, some individuals of r = 0.3125 or 0.25 share space to a greater degree than some individuals related by 0.5. Therefore, a close examination of the social structure and social dynamics of the population is necessary to explain the wide range in space-use overlap values. This examination will be presented in the discussion after describing patterns of space-use for different groups. Animals related by 0.5 were placed in three groups; mother:offspring, littermates, and nonlittermates. For each group, mean space-use overlap decreases as individuals become older (Fig. 2). However, littermates on average maintain a high overlap even into adulthood. Mother:offspring also maintain a high space-use overlap into adulthood whereas nonlittermate adults, although related by 0.5, have the lowest mean value, but still higher than the mean value for animals related by 0.25 or less (Fig 1). In all three groups, the range was greatest for adults. Similarly, spacing between individual full-sib hoary marmots (M. caligata) increases with age (Barash 1989: 215).
Animals related by 0.25 were placed in three groups; paternal half-sibs, grandmother:grand-offspring, and aunt:niece (Fig. 3). The highest mean space-use overlap (9.7%) occurred between adults and young (aunt:niece relationship) and this group also had the widest range of values. The range was the same as that of animals related by 0.3125 (Fig. 1); all of these individuals were nonlittermates that had the same father and were distantly related through their mothers. Space sharing by aunt:niece groups decreased from young to yearlings to adults. The mean space-use for the adult aunt:niece group (3.5%) was only slightly higher than that for unrelated adults (3.2%). Similarly, all groups of paternal half-sibs had low space-use overlap, especially when an adult or yearling was associated with a younger animal (Fig. 3). Again, the widest range was among adults. Finally, space-use overlap is low between grandmothers and their grandoffspring young.
In 1983, 855 became resident in Lower Picnic and by 1991 recruited six reproductive daughters (Armitage 1992). The seven females formed three matrilines (Table 1). Thus matriline one consisted of the matriarch and two nonlittermate sisters; matriline two consisted of a pair of littermate sisters, and matriline three consisted of two nonlittermate sisters. All of their offspring had the same father and all offspring were related to the adult females through their mothers; therefore r =0.3125. Within each matriline, young had high space-use overlap, thus accounting for the high end of the range of values (Fig. 1). However, young of different matrilines had low or no space-use overlap, thus accounting for the values at the low end of the range. Because there were more values for young between matrilines than within matrilines, the mean value was relatively low and lower than predicted from the "proportional distribution of benefits" hypothesis.
The same matrilineal formation accounts for the wide range of space-use overlap values when the adult is an aunt and the young are nieces (Fig. 3). Thus overlap was high between the young of 478 and their aunt 497 and between the young of 497 and their aunt 478. Conversely, space-use overlap was low between the young of females in one matriline and their aunts in the other matrilines. Because there were many more overlap comparisons between matrilines than within matrilines, mean space-use overlap was only 9.7%.
The same matrilineal formation exemplifies the wide range of space-use overlap values for mother:offspring adults and littermate adults. Thus, 855 had high space-use overlap with her daughters in matriline l, but very low values of overlap with her daughters in matrilines 2 and 3 (Table 1). Conversely, 761 had high overlap with her nonlittermate sister 100, but low overlap with her littermate sister 733.
The high overlap of adult:adult paternal half-sisters occurred at Upper Picnic. Littermate sisters occupied the area in 1980 and established a matriline. Subsequently, descendants three generations later had the same father, but different mothers (there was a distant and very low relatedness through great-grandmothers). Because they were in the same matriline, they shared space. Most adult paternal half-sibs live in different matrilines and space-use overlap is very low, hence the low mean value (Fig. 3).
The high space-use overlap of close-kin among yellow-bellied marmots is similar to the pattern of space-use by the family unit of the alpine marmot (M. marmota). The family shares space but there is little overlap with neighbouring families (Perrin et al., 1993). By contrast, there is little overlap in space-use by female woodchucks (Meier 1992). In effect, each individual woodchuck forms a matriline of one.
Two major conclusions are apparent. First, benefits are not distributed proportionately by kinship, but are concentrated on close kin, particularly those related by 0.5. Because the mean values for space-use overlap for kin related by 0.125 or less are lower than the mean value for unrelated animals, kin related by < 0.25 can be considered to be treated as if they were unrelated. Unrelated adults have higher values of space-use overlap when they are in conflict and one adult makes excursions into the home range of another and initiates agonistic behavior, which results in the dominant female displacing the subordinate female from part or all other home range (Armitage 1992).
Second, matrilineal organization is the social mechanism whereby female yellow-bellied marmots attempt to maximize inclusive fitness. Although it is not always clear why particular individuals form a matriline, membership in a matriline rarely includes animals related by <0.5. Most matrilines are mother:daughter associations with sister:sister groups forming about 22% of all matrilines (Armitage 1989). Familiarity with members of a matriline seems to be the basis for social bonding and under rare circumstances includes individuals related by 0.25, but never <0.25. All space-use overlap of individuals related by <0.25 occurs at the boundaries of the home ranges (Fig. 10, Table 5 in Armitage 1989). Matrilineal structure and function strongly indicate that yellow-bellied marmots attempt to maximize direct fitness and that indirect fitness benefits arise as a secondary consequence of direct fitness strategies.
*****
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |