1ère Journée d'Étude sur la Marmotte Alpine, Ramousse R. & Le Berre M. eds. : 37-48.
ELEMENTS DE DEMOGRAPHIE ET RYTHME D'ACTIVITE D'UNE COLONIE DE MARMOTTES
(Marmota marmota)
DANS LE PARC NATIONAL DES PYRENEES OCCIDENTALES.
Gérard NOGUE et Christian ARTHUR

Résumé
De 1982 à 1989, de nombreuses observations ont été faites sur la marmotte, espèce réintroduite dans le Parc National des Pyrénées Occidentales. Sont présentées ici les données relatives à l'activité printanière et à la démographie de quelques familles. La période d'activité débute en général fin mars-début avril, le comportement de sortie étant stabilisé à partir de la fin avril. En mai, le rythme d'activité est bimodal avec une période de sortie maximale entre 8h30 et 10h30 et une seconde période en fin d'après-midi entre 16 et 18 heures. Le taux d'accroissement des populations a varié entre 0,072 et 0,098 selon les années. Sur certains sites occupés depuis longtemps, le taux d'accroissement annuel a été plus faible, r = 0,035. Les immatures disparaissent des familles entre le printemps et l'été (53 % des individus disparaissant), la mortalité des immatures et juvéniles est forte pendant l'hiver (30 %) alors que la survie des juvéniles pendant l'été et l'automne semble bonne. La reproduction est relativement constante d'une année sur l'autre et entre familles (3,18 jeunes). Tous ces résultats sont discutés : 1) pour définir une méthode d'estimation d'abondance ; 2) au plan de la régulation démographique des populations de marmottes.
Mots-clé
Marmotte, Marmota marmota, rythme d'activité, démographie, Pyrénées, reproduction, période d'activité.
I. INTRODUCTION
La marmotte (Marmota marmota), disparue des Pyrénées à la fin du Pléistocène (Besson, 1971), a été réintroduite dans ce massif dès 1948, mais surtout à partir de 1969 à l'initiative du Parc National des Pyrénées Occidentales (Besson, 1971). A partir de quelques points d'introduction, l'espèce a colonisé l'ensemble de la zone centrale du Parc, s'étendant rapidement dans un territoire vierge où les pressions sélectives tant naturelles (compétition intraspécifique, parasites et maladies, prédation ...) qu'humaine (chasse et destruction) étaient soit absentes, soit fortement réduites. C'est dans ce contexte biologique particulier qu'un agent technique du Parc National des Pyrénées Occidentales (G. Nogué) a entrepris le suivi démographique et l'étude de la répartition de l'espèce, essentiellement dans la vallée où ont eu lieu les premières introductions. Nous présentons ici les premiers résultats concernant le rythme d'activité de quatre familles, regroupées en une colonie, qui ont été étudiées plus particulièrement de 1982 à 1989 ainsi que la démographie de ces familles et d'autres colonies présentés dans différentes vallées du secteur de Luz dans le PNPO. L'ensemble des données récoltées durant cette période seront reprises plus en détail lors de publications ultérieures.
II. ETUDE DU RYTHME D'ACTIVITE DE LA MARMOTTE DANS LES PYRENEES
A. Durée de la période d'activité
Méthode d'étude :
En 1982, 1986 et 1989 les terriers des différentes familles constituant la colonie du Pla de Bert ont été examinés tous les jours à partir du 20 mars pour la sortie printanière, et à partir du 10 octobre pour l'entrée en hibernation. Cette colonie est située au-dessus du hameau d'Héas, rive droite du cours d'eau des Touyères. Son territoire couvre une superficie de 23 hectares à l'altitude moyenne de 1650 mètres. Il est exposé au Nord-Ouest et sa formation est de deux tiers de pelouse pour un tiers d'éléments rocheux. Pour la sortie printanière, ont été notés le premier jour où un terrier était débouché et un trou de marmotte observé, ainsi que le premier jour où tous les terriers étaient débouchés. Pour l'entrée en hibernation, seul a été noté le dernier jour où un trou de marmotte était observé.
Résultats :
Pour la colonie étudiée, la période d'activité a été très constante pour les 3 années, soit un peu plus de 6 mois et demi, et ce en dépit de l'augmentation du nombre de familles constituant cette colonie, ainsi que du nombre d'individus (tableau 1).
| Année
| 1982 | 1986 | 1989
|
|---|
| Période | Sortie
d'hibernation | Entrée en
hibernation | Sortie | Entrée | Sortie | Entrée
|
|---|
| Date | 08 avril | 20 octobre | 02-05 avril | 28 octobre | 31 mars-03 avril | 23 octobre
|
|---|
| Durée
| 200 jours | 209 jours | 207 jours
|
|---|
| Nombre de familles différentes (nombre minimum d'individus)
| 4 (23) | 6 (38) | 7 (42)
|
|---|
Tableau 1. Durée de la période d'activité de la colonie de marmottes du Pla de Bert, Parc National des Pyrénées Occidentales (altitude 1650 m ; exposition au Nord-Ouest).
En plus d'un rythme endocrine propre à chaque colonie, l'exposition et l'altitude semblent intervenir sur la sortie d'hibernation, les colonies exposées au nord, ou se situant en altitude, ayant tendance à sortir plus tard, parfois au-delà du 20 avril. L'exposition peut aussi contrebalancer les effets de l'altitude, la colonie de l'Aguila, située à proximité de la colonie du Pla de Bert mais plus haut (1900 mètres) et exposée au Sud-ouest, ayant tendance à sortir d'hibernation plus tôt que cette dernière colonie (Vougny, 1990).
B. Etude du comportement de sortie printanier:
Méthode d'étude :
En 1989, sur une colonie de marmottes, la colonie du Pla de Bert, à partir du moment où les premières traces d'activité printanière ont été relevées, un dénombrement des individus présents au dehors a été effectué à intervalles plus ou moins réguliers. Le décompte a eu lieu soit entre 8 heures et 10 heures (heure légale), soit entre 16 et 18 heures. Deux balayages de la zone étaient faits à 15 minutes d'intervalle, les individus comptés lors de ces 2 balayages étant cumulés dans le décompte. Les dénombrements ont été arrâtés une fois le nombre d'animaux comptés stabilisé sur une longue période.
Résultats :
La figure 1 retrace pour la période allant du 01 avril au 23 mai 1989, le nombre de marmottes comptées à intervalles de 2-5 jours (hormis du 18 avril au 01 mai). On constate que ce nombre augmente lentement du 01 au 12 avril (fortes giboulées de neige le 12 avril expliquant la chute dans les comptages) puis rapidement jusqu'au 18 avril. A partir du 1er mai, le nombre de marmottes comptées à un quart d'heure d'intervalle est stabilisé et fluctue très peu ( de 35 à 40 individus) malgré les variations des conditions météorologiques.
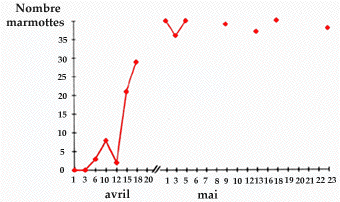 Figure 1. Etude du comportement de sortie printanière de la marmotte dans le PNPO.
Figure 1. Etude du comportement de sortie printanière de la marmotte dans le PNPO.
Evolution du nombre de marmottes sorties entre le 1er avril et le 23 mai 1989 sur la colonie du Pla de Bert (7 familles ; altitude 1650 m, exposition au nord-ouest).
Entre le 18 avril et le 1er mai, sur 10 passages auprès de la colonie, 5 ont eu lieu par temps de neige, brouillard et pluie ; aussi les nombres de marmottes comptées n'ont pas été pris en compte.
C. Etude du rythme d'activité journalier printanier
Méthodes d'étude et d'analyse :
Les 03 mai 1986 et 01 mai 1989, de 7 heures à 19 heures, heure légale (soit de 06 à 18 heures, heure solaire), toutes les marmottes visibles de 3 familles de la colonie du Pla de Bert (familles Mail de Bert, Caübe, Canal) ont été dénombrées par balayage à partir du mâme point d'observation. Le balayage, effectué toutes les quinze minutes, a toujours été fait dans le mâme sens. La météorologie a été sensiblement identique les deux jours : couvert le matin avec quelques flocons de neige, puis éclaircies et soleil à partir de 10 heures, les 3/4 du territoire étant déneigés pour le 03 mai 1986 ; beau temps frais, moitié du territoire déneigé le 01 mai 1989.
La comparaison des résultats entre années a été faite au moyen d'une analyse de variance à 3 facteurs (année, heure, pointage) sans repliquats, la variable étudiée étant le nombre de marmottes présentes au dehors et visibles (logiciel STATITCF, Phelippeau, 1985). Aucune transformation n'a été effectuée sur les variables, l'homoscédasticité étant vérifiée.
Résultats
La figure 2 retrace, pour les deux années, l'évolution du nombre de marmottes dénombrées heure par heure.
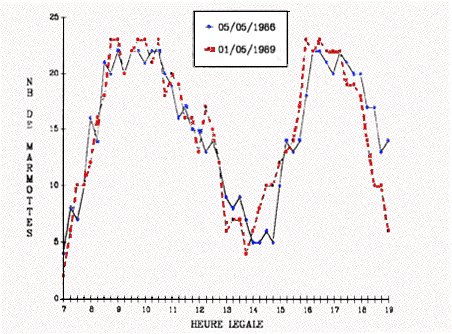 Figure 2. Etude du rythme d'activité journalier de la marmotte au printemps dans le PNPO.
Figure 2. Etude du rythme d'activité journalier de la marmotte au printemps dans le PNPO.
Evolution du nombre de marmottes comptées toutes les 15 minutes sur 3 familles de la colonie du Pla de Bert (altitude 1650 m ; exposition au Nord-Ouest).
On observe chaque année une courbe bimodale, les marmottes commençant à sortir dès 7 heures (06 heures, heure solaire) ; leur nombre se stabilise entre 8 heures 30 et 10 heures 30, période où un premier maximum est atteint (de 20 à 23 individus). A partir de 10 heures 30, le nombre d'individus observé diminue régulièrement jusqu'à atteindre un minimum entre 13 heures et 15 heures (entre 4 et 8 individus). Leur nombre remonte ensuite rapidement pour atteindre un second maximum (de 21 à 23 individus) entre 16 heures et 18 heures.
L'analyse de variance montre un comportement de sortie identique entre années (F année = 0,02 ; P = 0,889), une absence de variation significative entre pointages effectués durant une heure donnée (F pointage = 1,22 ; P = 0,316) et une variation très fortement significative du nombre de marmottes en fonction de l'heure d'observation (F heure = 272,66 ; P < 0,0001). Les trois facteurs (année, heure, pointage) expliquent par ailleurs 90,2 % de la variance observée dans les dénombrements.
D. Discussion - Conclusion
Notre intention n'est pas de discuter les résultats obtenus à la lumière des différents travaux conduits sur le comportement et le rythme d'activité de la marmotte alpine. Les Parcs Nationaux étant chargés, en ce qui concerne les espèces animales, d'une mission d'inventaire patrimonial, avec un suivi ultérieur de l'évolution des populations, les résultats acquis seront analysés dans l'optique de la définition d'une méthode de suivi des populations de marmottes par indice ponctuel d'abondance (I.P.A.). On remarquera cependant que le comportement printanier (notamment du réveil) décrit pour les Pyrénées est identique à celui observé par Zelenka (1965), les dates de sortie s'échelonnant dans les Alpes entre le début et la mi-avril. Les mâmes effets, parfois contradictoires, de l'altitude et de l'exposition sur le réveil ont aussi été notés par cet auteur.
Cette constance dans le cycle saisonnier, tous lieux confondus, et la permanence du rythme d'activité printanier observé tant dans les Alpes (Zelenka 1965) que dans les Pyrénées nous conduisent cependant à penser que la méthodologie proposée ci-dessous peut âtre appliquée sur toute l'aire de distribution géographique de la marmotte.
Les populations de marmottes peuvent âtre considérées comme étant à leur minimum après la sortie d'hibernation, les colonies ayant subi la mortalité hivernale et les jeunes n'étant pas encore nés. C'est cette période qui a été choisie pour le dénombrement des individus en vue de l'établissement d'un indice d'abondance.
L'examen des figures 1 et 2 et des résultats de l'analyse de variance permettent de définir une méthodologie facilement applicable à grande échelle, susceptible de fournir une bonne image de l'évolution des populations. Le comportement de sortie printanier étant stabilisé à partir de la fin du mois d'avril, nous disposons d'environ 1 mois (tout mai) pour échantillonner un maximum de colonies de marmottes. Cet échantillonnage peut s'effectuer sur la base de dénombrements à poste fixe des individus sortis des terriers. L'absence de variations significatives entre deux pointages effectués à un intervalle de 15 minutes pour une heure donnée permet de se restreindre à ce nombre (2 pointages) et donc durant un laps de temps donné d'échantillonner plusieurs colonies. L'échantillonnage est par contre à effectuer aux heures de sortie maximum des marmottes qui correspondent aussi aux périodes de plus forte stabilité comportementale (moins de 10 % de variation entre pointage durant ces périodes), soit entre 8 h 30 et 10 h 30 (heure légale) le matin ou entre 16 h 00 et 18 h 00 le soir (heure légale). En considérant un déplacement d'environ 15 minutes entre 2 colonies et un temps de latence de quinze minutes entre l'arrivée sur une colonie et le premier pointage, il serait alors possible d'échantillonner 6 colonies par jour sur une vallée. Cinq à six vallées pouvant âtre prospectées par secteur du Parc au cours du mois de mai, on obtiendrait alors un total de près de 250 colonies échantillonnées (6 colonies/jour sur 6 vallées pendant 7 jours) sur l'ensemble de la zone centrale du PNPO. Cet échantillonnage, associé à un repérage des colonies au moment de leurs premières sorties annuelles (les terriers débouchés et les traces des animaux sont très visibles sur la neige fin mars, début avril), permettrait tous les trois ans d'obtenir une bonne image de la répartition et de l'abondance en marmottes dans la zone centrale du PNPO. Il resterait alors à effectuer une stratification des colonies échantillonnées en fonction de l'altitude, de l'exposition et des types d'habitats pour optimiser la précision des IPA. Le dernier élément à vérifier consisterait à étudier la relation entre le nombre de marmottes comptées et le nombre réellement présent pour obtenir des valeurs précises de densité.
III. ELEMENTS DE DEMOGRAPHIE DE LA MARMOTTE DANS LES PYRENEES
A. Evolution démographique dans trois vallées
Méthode d'étude
Dans trois vallées du PNPO, Estaubé, Héas et Campbieil (secteur de Luz-St-Sauveur), les colonies de marmottes ont été répertoriées et le nombre d'individus les fréquentant estimé par observation directe entre 20 et 35 jours après le réveil printanier, le décompte étant effectué entre 8 h 00 et 11 h 00 du matin et 15 h 00 et 18 h 00 le soir (heure légale). A l'intérieur de ces trois vallées, la zone étudiée couvre une superficie de 5 000 hectares environ. Son altitude varie de 1300 à 3000 mètres. Formée essentiellement de pelouses rases, d'éboulis et de parois rocheuses, elle est fréquentée, outre la marmotte, par l'isard, le lièvre d'Europe, les vaches, les moutons et aussi des prédateurs tels que l'aigle royal et le renard.
Les dénombrements ont été effectués aux printemps 1982, 1986 et 1989. Chaque année les nouvelles colonies ont été recherchées et incluses dans le dénombrement. Toutes les colonies présentes en 1982 n'ont cependant pas été dénombrées cette année-là. Les facteurs année et vallée ont été comparés au moyen d'une analyse de variance.
Résultats
La figure 3 retrace pour les trois vallées et les trois années de dénombrement l'évolution du nombre moyen de marmottes par colonie. On observe une augmentation régulière du nombre qui, toutes vallées confondues, passe de 8,16 marmottes/colonie en 1982 à 11,37 en 1986 et 13,84 en 1989. L'augmentation la plus spectaculaire est notée entre 1982 et 1986 en vallée d'Estaubé où le nombre moyen de marmottes/colonie passe de 5,83 à 10,8. Globalement cependant aucune différence significative dans l'abondance en marmottes/colonie n'apparaît entre vallées (F ajusté = 0,999 ; P > 0,05) ou entre année (F ajusté = 4,637 ; P>0,05) ce qui indique une augmentation régulière et continue sur les 7 années d'étude tant entre années qu'entre vallées. La variance résiduelle est toutefois encore forte (18 %) ce qui laisse supposer des évolutions démographiques variables selon les colonies. On remarque que l'abondance toutes colonies en 86 et 89 en vallée de Campbieil est inférieure à celle des 5 colonies comptées régulièrement en 82, 86 et 89, ce qui semble indiquer que certaines colonies n'avaient pas été comptées en 82 car présentant une faible abondance.
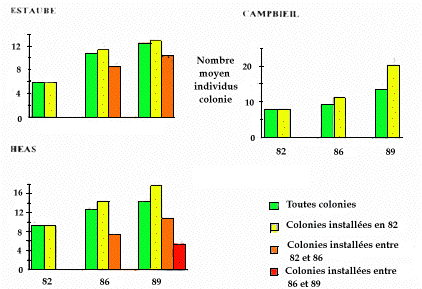 Figure 3. Evolution du nombre moyen de marmottes comptées par colonie en 1982, 1986 et 1989 dans trois vallées du PNPO (n = nombre de colonies dénombrées).
Figure 3. Evolution du nombre moyen de marmottes comptées par colonie en 1982, 1986 et 1989 dans trois vallées du PNPO (n = nombre de colonies dénombrées).
(Nota = la différence entre le nombre total de colonies présentes et la somme des colonies dénombrées en 82, 86 et 89 et les nouvelles colonies installées en 86 et 89 vient du fait que toutes les colonies présentes en 82 n'ont pas été recensées cette année-là).
Colonie par colonie, certaines "anomalies" peuvent âtre constatées ; ainsi la colonie n° 1 en vallée d'Estaubé qui passe de 5 individus en 1982 à 24 en 1986, ou encore la colonie n° 19 en vallée d'Héas qui passe de 19 à 32 individus de 1982 à 1986 ou la colonie n° 7 en vallée de Campbieil qui évolue de 18 individus en 1986 à 40 individus en 1989.
Globalement, toutes vallées et colonies confondues, le taux moyen d'accroissement annuel, r, est égal à 0,098 entre 1982 et 1986 et à 0,072 entre 1986 et 1989 (voir Lebreton 1981 pour le mode de calcul de r).
Entre 1982 et 1986, les colonies anciennement installées ont connu, toutes vallées confondues, un taux d'accroissement moyen annuel égal à 0,178, ce taux chutant à 0,073 entre 1986 et 1989, alors que les colonies nouvellement installées en 1986 ont connu entre 1986 et 1989 un taux d'accroissement moyen annuel de 0,136.
B. Evolution démographique de la colonie du Pla de Bert
Méthode d'étude
De 1982 à 1989, la colonie du Pla de Bert a été recensée chaque année au printemps selon la méthode décrite au § III. A. Le nombre de familles présentes a été estimé après report sur une carte des observations des animaux chaque printemps.
Résultats
La figure 4 retrace de 1982 à 1989 (hormis 1987 et 1988) le nombre de familles présentes et le nombre total d'individus recensés au printemps (adultes + immatures) chaque année. On remarque que l'accroissement est faible de 1982 à 1984 (r = 0,042 ; aucune nouvelle famille), fort de 1984 à 1985 (r = 0,32 ; 2 nouvelles familles), moyen entre 1985 et 1986 (r = 0,152 ; aucune nouvelle famille) et à nouveau faible entre 1986 et 1989 (r = 0,035 ; 1 nouvelle famille).
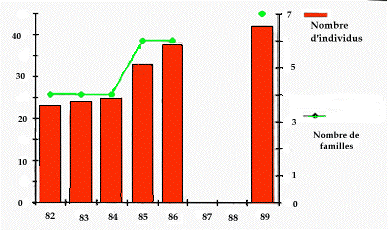 Figure 4. Evolution du nombre de familles et du nombre total d'individus (adultes + immatures) de la colonie du Pla de Bert, PNPO de 1982 à 1989 (pas de recensement en 1987 et 1988).
Figure 4. Evolution du nombre de familles et du nombre total d'individus (adultes + immatures) de la colonie du Pla de Bert, PNPO de 1982 à 1989 (pas de recensement en 1987 et 1988).
Comparativement aux taux d'accroissement observé sur l'ensemble des 3 vallées, le taux d'accroissement de la colonie du Pla de Bert est plus faible tant entre 1982 et 1986 (r moyen = 0,041 versus 0,098) qu'entre 1986 et 1989 (r moyen = 0,035 versus 0,073).
C. Evolution démographique sur deux années de familles de marmottes
Méthode d'étude
En 1982 et 1983, 4 familles de la colonie du Pla de Bert ont été recensées par observation directe aux trois saisons principales : printemps en mai, été en juin-juillet (époque de naissance et sortie des marmottons) et automne en septembre-octobre (avant l'entrée en hibernation). L'examen attentif des individus et leur localisation précise a permis de les rattacher à une famille et à une classe d'âge, malgré l'absence de marquage individuel. Trois classes d'âge ont été distinguées : adultes, immatures (individus nés l'année d'avant) et juvéniles (animaux nés dans l'année).
Les observations sont effectuées avec des jumelles 15 x 60 et une longue-vue grossissement 20-60 à une distance variant de 20 à 300 mètres.
En 1983, cinq marmottes ont été capturées à l'aide de pièges à mâchoires placés à l'entrée des terriers au printemps dans ces quatre familles. Marquées à l'oreille et équipées d'un collier de matière plastique colorée, elles ont été observées pendant environ 2,5 mois.
Résultats
Le tableau 2 retrace pour les quatre familles l'évolution du nombre d'individus par saison pour chaque classe d'âge. A partir de ces éléments nous avons calculé un taux de disparition (selon la méthode de Mann et Janeau, 1988) pour les adultes et immatures du printemps 1982 au printemps 1983, pour les jeunes de l'été 1982 au printemps 1983 (ils sont alors immatures) et pour les immatures entre le printemps et l'été en 1982 et 1983.
| | Anné et saison
|
|---|
| | 1982 | 1983
|
|---|
| Famille | Classe d'âge | P | E | A | P | E | A
|
|---|
| Mail Bernes | AD
IM
JUV
IND | 2
2
-
1 | 2
1
3
- | 2
1
3
1 | 3
2
-
1 | 2
1
2
- | -
-
-
-
|
|---|
| Coste de Bert | AD
IM
JUV
IND | 2
2
-
- | 2
-
2
- | 2
-
2
- | 2
2
-
- | 2
1
3
- | -
-
-
-
|
|---|
| Mail de Bert | AD
IM
JUV
IND | 2
3
-
- | 2
1
4
- | 2
1
4
1 | 3 (1 F)
2
-
1 | 2
1
3
1 | -
-
-
-
|
|---|
| Caube | AD
IM
JUV
IND | 4
5
-
- | 3
2
4
- | 3
2
4
2 | 4 (2 M)
3 (2 M)
-
1 | 3
2
3
1 | -
-
-
- |
|---|
Tableau 2. Composition par classe d'âge de quatre familles de marmottes de la colonie du Pla de Bert aux différentes saisons 1982 et 1983 dans le PNPO (sexe des animaux capturés en 1983).
La figure 5 retrace les résultats obtenus. On observe une faible disparition des adultes entre le printemps et l'été (18 % en moyenne), une forte disparition des immatures entre le printemps et l'été (53 % en moyenne), très différente entre 1982 (- 67 %) et 1983 (- 44 %), et une bonne survie des jeunes de l'été à l'automne suivie d'une disparition prononcée (- 31 %) en hiver.
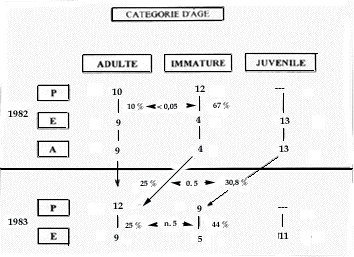 Figure 5. Taux de disparition des animaux en fonction de la catégorie d'âge et de la saison sur quatre familles de la colonie du Pla de Bert, PNPO.
Figure 5. Taux de disparition des animaux en fonction de la catégorie d'âge et de la saison sur quatre familles de la colonie du Pla de Bert, PNPO.
(Nota : les indéterminés n'ont pas été pris en compte).
D. Evolution annuelle de la reproduction de familles de marmottes
Méthode d'étude
De 1982 à 1989, chaque année (hormis 1987) le nombre de jeunes marmottes présentes dans les quatre familles étudiées ci-dessus a été estimé au début de l'été par observation. Les jeunes marmottes ont été rattachées à une famille sur la base de leur répartition et de leurs contacts avec les adultes.
Résultats
Ils sont reportés au tableau 3.
| Année
|
| Famille | 1982 | 1983 | 1984 | 1985 | 1986 | 1988 | 1989
|
|---|
| Mail Bernes | 3 (5) | 2 (6) | 2 (6) | 3 (5) | 2 (6) | 2 (?) | 4 (6)
|
|---|
| Coste de Bert | 2 (4) | 3 (4) | 3 (3) | 3 (5) | 2 (5) | 3 (?) | 1 (4)
|
|---|
| Mail de Bert | 4 (5) | 3 (6) | 4 (7) | 1 (7) | 3 (7) | 4 (?) | 4 (7)
|
|---|
| Caube | 4 (9) | 3 (8) | 4 8) | 5 (12) | 3 (10) | 4 (?) | 5 (9)
|
|---|
Tableau 3. Nombre de jeunes marmottes par famille pour quatre familles de la colonie du Pla de Bert, PNPO de 1982 à 1989 (nombre d'adultes et immatures présents dans la famille au printemps).
Globalement, toutes familles et années confondues, nous obtenons un nombre moyen de 3,18 > 1,09 jeunes/famille (sur 28 années-famille). Famille par famille, le nombre moyen de jeunes varie sur les 7 années de 2,43 jeunes pour la famille de Coste de Bert à 4.0 jeunes pour la famille de Caübe. Année par année, le nombre moyen de jeunes/famille varie de 2,5 (en 1986) à 3,5 (en 1989), ce nombre pouvant osciller pour une famille et une année donnée entre 1 et 5 jeunes. Le nombre moyen de jeunes/famille est plus ou moins fonction du nombre d'adultes présents au printemps, la corrélation entre les 2 nombres s'avérant positive (nombre de jeunes/famille = 1,10 (nombre d'adultes dans la famille) + 3,13 et significative (Cr = 0,55 ; P < 0,01).
E. Discussion
Comparativement aux résultats de Mann et Janeau (1988) dans les Alpes de 1976 à 1987, les populations de marmottes dans le PNPO ont présenté une dynamique d'accroissement continue de 1982 à 1989. Le taux d'accroissement moyen annuel (entre 0.07 et 0.10) est cependant nettement inférieur au taux potentiel théorique avancé par Huber (in traduction ONC 1978), qui est estimé à 0,15-0,30. Ce taux d'accroissement est très variable selon les familles, alors qu'il varie peu d'une vallée à l'autre et entre années. "L'âge" des colonies intervient pour moduler ce taux d'accroissement, les colonies nouvellement installées présentant, dans les années suivant leur création, un taux d'accroissement supérieur à celui des anciennes colonies.
Le nombre moyen de jeunes par famille est dans les Pyrénées supérieur à celui observé par Mann et Janeau (2,3) dans les Alpes. Ce nombre varie peu selon les années et est, semble t-il, influencé pour partie par le nombre d'adultes présents au printemps. Les différents auteurs ayant travaillé sur la marmotte mentionnant que seule la femelle adulte du groupe familial se reproduit, une telle corrélation (nombre de jeunes - nombre d'adultes) ne devrait pas âtre observée. Ce qui pose le problème de la validité de la définition des familles observées. Il est donc possible que certaines familles que nous avons observées soient en fait le regroupement de 2 familles, d'où une surestimation de la reproduction par famille.
Le schéma de fonctionnement démographique suivant pourrait in fine âtre proposé pour la marmotte dans les Pyrénées : fort accroissement des familles ; éclatement des familles avec constitution de deux groupes familiaux par division des territoires familiaux ; diminution de l'accroissement des groupes familiaux par ralentissement de la production de jeunes (moins de jeunes et/ou intervalle supérieur entre deux gestations) ; prospection estivale de nouveaux territoires par des adultes inappariés et/ou des immatures ; dispersion des adultes inappariés et immatures sous la pression sociale, qui s'installent définitivement sur les nouveaux territoires : fort accroissement sur ces nouveaux territoires, accroissement ralenti sur les anciens territoires.
Un tel fonctionnement, qui implique une forte influence de la régulation sociale, se ferait par à coups et expliquerait la dynamique contrastée observée entre 1982-1986 et 1986-1989. Il serait encore possible du fait de l'existence de zones potentielles encore non colonisées dans les Pyrénées. Les prévisions de l'expansion ultérieure de la marmotte dans les Pyrénées nécessiteraient alors une description fine des caractéristiques écologiques de l'habitat de l'espèce et une cartographie des zones restantes à coloniser.
IV. BIBLIOGRAPHIE
BESSON J.P. (1971). Introduction de la marmotte dans les Pyrénées Occidentales. C.R. du 96ème Congrès des Sociétés Savantes, Toulouse, 3 : 397-399.
HUBER W. (traduction ONC 1978). La marmotte des Alpes. O.N.C. Paris, 36 pp.
LEBRETON J. D. (1981). Contribution à la dynamique des populations d'oiseaux. Modèles mathématiques en temps discret. Thèse d'Etat, Université de Lyon, Claude Bernard, Villeurbanne, France : 253pp.
MANN C., JANEAU G. (1988). Occupation de l'espace, structure sociale et dynamique d'une population de marmottes des Alpes (Marmota marmota L.). Gibier Faune Sauvage, 5 : 427-446.
PHELIPPEAU P. (1985). Logiciel de statistiques, STATITCF. Comment interpréter les résultats d'une analyse de variance. I.T.C.F. Paris : 52pp.
VOUGNY P. (1990). Capture et marquage de la marmotte dans les Pyrénées Occidentales. Rapport de stage, E.N.V. Toulouse-P.N.P.O.: 12 pp.
ZELENKA G. (1965). Observation sur l'écologie de la marmotte des Alpes. La Terre et la Vie, 3 : 238-256.
Collaboration
Laboratoire Départemental des Services Vétérinaires 65 (pour autopsies)
Cadre administratif
Opération entièrement prise en charge par le Parc National des Pyrénées.
A suivre...
retour ou sommaire