
Social dynamics, Kinship, and Population Dynamics of marmot
ARMITAGE K.B.
Departement of Systematics and Ecology,
The University of Kansas, Lawrence, KS 66045-2106
USA
Probably the social systems of all marmot species, except
M. monax are based on kinship and most species have a family with a dominant breeding pair
and non-breeding subordinates. Social behavior of closely-related animals is primarily
amicable whereas agonistic behavior characterizes the social interactions of more
distantly-related or unrelated individuals. All social groups are characterized by reproductive
suppression of younger/subordinate animals by older/dominant individuals. Only the
solitary
M. monax lacks reproductive suppression among adults. Population growth occurs when offspring
are retained in their natal group. Recruitment of offspring occurs when they are
treated cohesively; dispersal is associated with either a lack of cohesive behaviors
or the presence of agonistic behaviors. Population saturation probably occurs in all species
and the lack of nutritious food is proposed as the limiting factor. Major population
decline usually is associated with unusual weather; e. g. drought or prolonged winter. The availability of wide-spread habitat provided ecological release that permitted
M. monax and
M. flaviventris to evolve different social systems as a mechanism for escaping reproductive suppression.
Key-words
: Marmots, Reproductive suppression, kinship, ecological release, social dynamics.
| Social system | Comments | Species | References |
| Solitary | Male pobably polygynous. Little overlap of female home ranges. | M. monax | Meier 1992 |
| Monogamous | Family groups with dominant reproductive pair. Non reproductive adults frequently present. Little home range overlap with neighboring families. | M. marmota | Perrin et al
1993 Arnold 1990 Mann & Janeau 1988 |
| Colonies consist of two or more family groups. Probably characteristic of all the Eurasian species. Polyandry possible in those species in which two or more adult males are present | M. baibacina
M. menzbieri M. bobac
M. sibirica
M. broweri (?) | Bibikov 1995 and references therein Rausch & Rausch 1971 | |
| Most often monogamous. | M. vancouverensis |
Bryant 1994 | |
| Polygynous | An adult male, two adult females, and a litter of yearlings and/or young occasionally some 2-yr-olds. | M. olympus | Barash 1973 |
| An adult male, two or three adult females, two or three 2-yr-olds, yearlings and young. | M. caligata | Barash 1974a Holmes 1984a | |
| Some populations monogamous An adult male associated with one or more matrilines of related females. | M. flaviventris | Armitage 1994 |
| Species | Young | Yearling | Young & Yearling | 2-yr-old | 3 or older | References |
| M. caudata | 46-70 (young, yearling & 2-yr-old) | 30-54 | Kiziliov & Berendyaev 1978* | |||
| M. baibacina | ||||||
| Mongolia (ave. of 4 yrs) | 45 | 11 | 44 | Bibikov 1995 | ||
| saturated population | 39 | 10 | 10 | 41 | ||
| Tien Shan | 35 | 13 | 52 | Bibikov 1967* | ||
| M. bobac shaganensis | 35 | 28 | 38 | Shubin 1988* | ||
| M. menzbieri | 26 | 29 | 45 | Mashkin 1981* | ||
| M. vancouverensis | ||||||
| natural colony | 14 | 7 | 29 | 50 | Bryant 1994 | |
| M. marmota | 28 | 21 | 24 | 28 | Perrin et al. 1993a | |
| 33 | 12 | 12 | 43 | Sala et al. 1992 | ||
| 25 | 21 | 14 | 41 | Arnold 1993 | ||
| M. olympus | ||||||
| (3-year average) | 31 | 24 | 13 | 32 | Barash 1973 | |
| M. flaviventris | ||||||
| (32-year average) | 37 | 22 | 45 | Schwartz et al . 1994 | ||
| M. monax | 39 | 61 (yearlings & adults) | Snyder 1962 | |||
| 48 | 20 | 31 (aged 2 or older) | Nuckle & Bergeron 1983 | |||
| Species | Age of First Reproduction | Percentage of adult females that breed | Reference |
| M. monax | 1 | 72 | Snyder & Christian 1960 |
| M. baibacina | 2 | 22 | Chervyakova & Bibikov 1971*, Bibikov 1989* |
| 2 | 33 | Bibikov 1967* | |
| M. marmota | 2 | 36 | Sala et al. 1992, Perrin et al . 1993a, Mann et al. 1993 |
| M. vancouverensis | 3 | 53 | Bryant 1994 |
| M. olympus | 3 | 41 | Barash 1973 |
| M. caligata | 3 | 43 | Barash 1989 |
| M. flaviventris | 2 | 52 | Armitage 1991 |
| 3% of 2-yr-old, 20% of 3-yr-old, 45% of 4-yr-old, and up to 70% of older females breed | Chervyakova & Bibikov 1971* |
| 3% of 2-yr-old, 18% of 3-yr-old, 39% of 4-yr-old, and 54% of aged 5 or older breed | Bibikov 1967* |
| reduced proportion of breeding females with increased population density, 75% at low to 47% at high | Pole 1992 |
| increased breeding in 2-yr-old animals in reduced population Pole & | Bibikov 1991* |
| decreased litter size with increased density | Mikhailyuta 1983*
|
| none of 47 animals under age 3 bred | Bibikov 1995
|
| proportion of breeding females and litter size decline at high density, about 25% of 2-yr-old animals breed at low density | Bibikov 1991*
|
| non breeding subadults | Mann & Janeau 1988 |
| non breeding subordinate adults | Arnold 1990
|
| most females do not breed until age 4 or older (x = 4.38 yrs) | Bryant 1994
|
| reproductive skipping when co-resident breeds, subordinate females produce half as many young as dominant females Wasser & | Barash 1983
|
| lowered reproduction in 2-yr-old females when adult females, including their mothers, are present | Armitage 1989 |
| weaning succes lower in females living in proximity to other adult females than in those living solitarily, reduced litter production in larger harems | Armitage 1986a
|
| more yearling females breed at low population density | Snyder 1962 |
Social systems of marmots
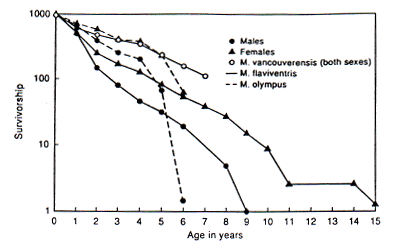
Social systems and survivorship
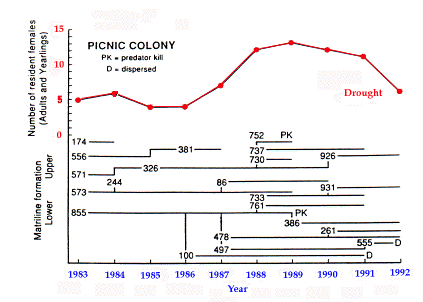
Reproductive suppression
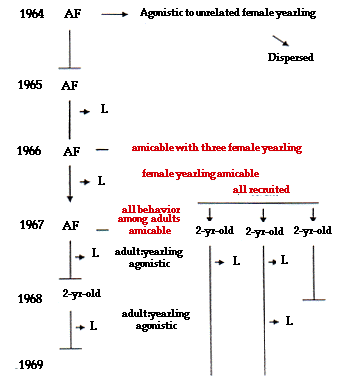
Social behavior and kinship
In general, monogamous family groups are characterized by a lack of agonistic behavior,
which occurs mainly when family units are disrupted (Bibikov 1995). In
M. menzbieri, greetings and groomings predominate within the family group and agonistic behavior
is directed toward intruders at the edge of the family range (Mashkin 1983, cited
in Bibikov 1995). In the Kazakhstan bobac (
M. bobac), 87% of the social contacts were greetings and groomings, agonistic contacts occurred
at the boundaries of family territories between adult resident males (Shubin 1988,
cited in Bibikov 1995). Among
M. marmota, agonistic behavior was much less frequent than cohesive behavior and mostly involved
unfamiliar adult males. Agonistic behavior was more likely to occur between members
of different groups (Perrin
et al. 1993b).
Behavior, Kinship and population dynamics
The saturation hypothesis
Ecological release
Alexander R. D. 1974. The evolution of social behavior. Annu. Rev. Ecol. Syst.,
5: 325-383.
Anthony M. 1962. Activity and behavior of the woodchuck in southern Illinois. Occas. Papers Adams Ctr. Ecol. Studies,
6: 1-25.
Armitage K. B. 1973. Population changes and social behavior following colonization
by the yellow-bellied marmot. J. Mamm.,
54: 842-854.
Armitage K. B. 1975. Social behavior and population dynamics of marmots. Oikos,
26: 34 l-354.
Armitage K. B. 198l. Sociality as a life history tactic of ground squirrels. Oecologia
, 48: 36-49.
Armitage K. B. 1984. Recruitment in yellow-bellied marmot populations: kinship, philopatry,
and individual variability. In Biology of Ground-Dwelling Squirrels
, J. O. Murie & G. R.Michener, eds., Univ. Nebraska Press, Lincoln: 377-403
Armitage K. B. 1986a. Marmot polygyny revisited: determinants of male and female reproductive strategies. In Ecological Aspects of social Evolution
, D. S. Rubenstein & R. W. Wrangham, eds., Princeton Univ. Press, Princeton, NJ.: 303-331.
Armitage K. B. 1986b. Individuality, social behavior, and reproductive success in
yellow-bellied marmots. Ecology,
67: 1186-1193.
Armitage, K. B. 1988. Resources and social organization of ground-dwelling squirrels.
In The Ecology of social Behavior
, C. N. SIobodchikoff, ed., Academic Press, NY.: 131-155
Armitage K. B. 1989. The function of kin discrimination. Ethol. Ecol. Evol.,
1: 111-121.
Armitag, K. B. 199l. Social and population dynamics of yellow-bellied marmots: results
from long-term research. Annu. Rev. Ecol. Syst.
, 22: 379-407.
Armitage K. B. 1992. Social organization and fitness strategies of marmots. Proc. lst Int. Symp. on Alpine Marmot, Bassano B., Durio P., Gallo Orsi U., Macchi E., eds.: 89-94.
Armitage K. B. & D. W. Johns 1982. Kinship, reproductive strategies and social dynamics of yellow-bellied marmots. Behav. Ecol. Sociobiol
., 11: 55-63.
Arnold W. 1990. The evolution of marmot sociality: II. Costs and benefits of joint
hibernation. Behav. Ecol. Sociobiol
., 27: 239-246.
Arnold W. 1992. Social evolution and obligatory group hibernation in marmots. Proc. lst Int. Symp. on Alpine Marmot
, Bassano B., Durio P., Gallo Orsi U., Macchi E., eds.: 4l-52.
Arnold W. 1993. Social evolution in marmots and the adaptive value of joint hibernation.
Verh. Dtsch. Zool. Ges.
, 86: 79-93.
Barash D. P. 1973. The social biology of the Olympic marmot. An. Behav. Monogr
., 6: 171-245.
Barash D. P. 1974a. The social behaviour of the hoary marmot (Marmota caligata). Anim. Behav
., 221: 256-261.
Barash D. P. 1974b. Mother-infant relations in captive woodchucks (Marmota monax
). Anim. Behav., 22: 446-448.
Barash D. P. 1989. Marmots Social Behavior and Ecology
. Stanford Univ. Press, Stanford, CA.
Bibikov D. 1995. Marmots of the World
.
Brody A. K. & K. B. Armitage 1985. The effects of adult removal on dispersal of yearling
yellow-bellied marmots. Can. J. Zool.
, 63: 2560-2564.
Bryant A. A. 1994. Notes on the Vancouver Island marmot (Marmota vancouverensis). Vancouver Island Marmot Recovery Project Review Paper, No. 3: l-25, Unpublished.
Downhower J. F. & K. B. Armitage 198l. Dispersal of yearling yellow-bellied marmots
(Marmota flaviventris). Anim. Behav
., 291: 1064-1069.
Hamilton W. D. 1964. The genetical evolution of social behaviour. J. Theoret. Biol., 7: 1-52.
Holmes W. G. 1984. The ecological basis of monogamy in Alaskan hoary marmots. In Biology of Ground-Dwelling Squirrels, J. O. Murie & G. R. Michener, eds., Univ. Nebraska Press, Lincoln: .250-274
Johns D. W. & K. B. Armitage 1979. Behavioral ecology of alpine yellow-bellied marmots. Behav. Ecol. Sociobiol
., 5: 133-157.
Kilgore D. L. Jr. & K. B. Armitage 1978. Energetics of yellow-bellied marmot populations.
Ecology
, 59: 78-88.
Mann C. S. & G. Janeau 1988. Occupation de l'espace, structure sociale, et dynamique
d'une population de marmottes des Alpes (Marmota marmota
L.). Gibier Faune Sauvage, 5: 427-445.
Mann C. S., E. Macchi & G. Janeau 1993. Alpine marmot (Marmota marmota
L.). Ibex, J. M. E., 1: 17-30.
Meier P. T. 1992. Social organization of woodchucks (Marmota monax
). Behav. Ecol. Sociobiol
., 31: 393-400.
Nuckle J. R. & J.-M. Bergeron 1983. Choix d'une méthode d'estimation de densité de
population de la marmotte commune (Marmota monax
). Can. J. Zool.
, 61: 2476-2481.
Perrin C., J. Coulon & M. Le Berre 1993a. Social behavior of alpine marmots (Marmota marmota
): seasonal, group and individual variability. Can. J. Zool
., 71: 1945-1953.
Perrin C., D. Allaine & M. Le Berre 1993b. Socio-spatial organization and activity
distribution of the alpine marmot Marmota marmota
: preliminary results. Ethology
, 93: 21-30.
Pole S. B. 1992. Dynamics of the grey marmot population structure and size and the
mechanisms of their restoration in the Tien-Shan natural plague focus. Proc. lst Int. Symp. on Alpine Marmot
, Bassano B., Durio P., Gallo Orsi U., Macchi E., eds: 129-133.
Rausch R. L. & V. R. Rausch 197l. The somatic chromosomes of some North American marmots
(Sciuridae), with remarks on the relationships of Marmota broweri
Hall and Gilmore. Mammalia
, 35: 85-101.
Sala L., C. Sola A. Spampanato & P. Tongiorgi 1992. The marmot population of the Tuscan-Emilian Apennine Ridge. Proc. lst Int. Symp. on Alpine Marmot
, Bassano B., Durio P., Gallo Orsi U., Macchi E., eds.: 143-149.
Schwartz O. A., K. B. Armitage & D. Van Vuren 1994. A 32-year demography of a yellow-bellied
marmot population. Ibex J.M.E, 2:43..
Snyder R. L. 1962. Reproductive performance of a population of woodchucks after a
change in sex ratio. Ecology, 43: 506-515.
Snyder R. L. & J. J. Christian 1960. Reproductive cycle and litter size of the woodchuck. Ecology
, 41: 647-656.
Svendsen G. E. 1974. Behavioral and environmental factors in the spatial distribution
and population dynamics of a yellow-bellied marmot population. Ecology
, 55: 760-771 .
Van Vuren D. 1990. Dispersal of yellow-bellied marmots.
Ph.D. dissertation, Univ. Kansas, Lawrence.
Van Vuren D. & K. B. Armitage 1994a. Survival of dispersing and philopatric yellow-bellied marmots: what is the cost of dispersal? Oikos
, 69: 179-18l.
Van Vuren D. & K. B. Armitage 1994b. Reproductive success of colonial and noncolonial female yellow-bellied marmots (Marmota flaviventris). J. Mamm
., 75: 950-955.
Wasser A. K. & D. P. Barash 1983. Reproductive suppression among female mammals: implications for biomedicine and sexual selection theory. Q. Rev. Biol.
, 58: 513-538.
White T. C. R. 1993. The Inadequate Environment. Springer-Verlag, Berlin.
Williams G. E. 1966. Adaptation and Natural Selection.
Princeton Univ. Press, Princeton, NJ.
Wynne-Edwards V. C. 1962. Animal Dispersion in Relation to Social Behaviour
. OIiver & Boyd, Edinburgh.
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |