![]()
Les activités mensuelles d'une population de marmottes alpines
(Marmota marmota), située dans les pré-Alpes Juliennes,
ont été observées et analysées pour tester les
facteurs internes et externes pouvant influer sur leur comportement. Les
données ont été recueillies deux années
successives. L'organisation sociale et la démographie ont
été aussi décrites. Une tendance estivale de la
distribution des activités est discutée. Parmi les facteurs
externes, les conditions météorologiques semblent jouer un
rôle déterminant. L'hypothèse de Türk & Arnold
(1988) sur la fonction du repos chez la marmotte alpine est soutenue par nos
données. L'hypothèse d'une relation entre défense
territoriale et probabilité de rencontre des individus dispersants est
formulée.
Mots-clés : Marmota marmota, pré-Alpes Juliennes,
comportement.
Monthly activities of a population of alpine marmots (Marmota
marmota), living in the Julian Pre-Alps (NE Italy), have been observed and
analysed in order to investigate external and internal factors that may
influence their behaviour. Behavioural data have been collected for two
consecutive years. Social organisation and demography are also described. A
summer trend in the distribution of activities is discussed. Among the external
factors, meteorological conditions seem to play a major role. Turk and Arnold's
(1988) hypothesis about the function of lying in alpine marmot is supported by
our data. An hypothesis relating territory defence and probability of meeting
dispersing individuals is formulated.
Key words: Alpine marmot, Marmota marmota, Julian Pre-Alps, behaviour.

La marmotte alpine (Marmota marmota) vit au sommet des montagnes, dans un environnement extrême à fortes variations météorologiques journalières et sai-sonnières. Pour y survivre, la marmotte a développé des adaptations physiologiques et comportementales. Le but de ce travail est d'étudier, au travers de l'activité men-suelle de M. marmota, certains des facteurs internes et externes qui pourraient influer sur son comportement.
Méthodes d'échantillonnage : La population étudiée, non autochtone, était composée de 2 familles contiguës (A et B). Le nombre maximum d'individus observé a été de 18. Le recueil des données a duré deux saisons (1990-91), de mars à octobre, plus deux mois en 1992; soit 51 jours ou 568 heures. Les animaux ont été observés à la jumelle (7x50 et 8x35) et au télescope (30x100) par au moins deux observateurs en même temps. Les méthodes d'échantillonnage par balayage et par focalisation ont été employées (Altmann 1974). Les comportements ont été enregistrés suivant un étho-gramme fourni gracieusement par le Dr. Lenti-Boero (non publié). Les animaux n'ont pas été marqués. Les données ont été regroupées, selon Lehner (1979), en états (affouragement, postures, locomotion, récolte et transport d'herbe) et en événements (salutations, inter-actions de jeu, auto et hétéro-toilettage, marquages de patte ou de joue, interactions agonistiques). Les com-portements actifs (affouragement, locomotion, récolte et transport d'herbes et jeu) et non-actifs (postures assise ou couchée) ont été distingués. Les animaux non visi-bles ont été considérés comme étant dans les terriers.
Analyse des données : les données comporte-mentales et météorologiques recueillies sur le terrain ont été codées et enregistrées dans une base de données. Quelques analyses ont été réalisées grâce au logiciel statistique SPSS, et les autres grâce à 10 programmes originaux en Fortran 77. Le coefficient de Spearman (rs) a été utilisé pour les corrélations.
Au cours de quatre années, la famille A s'est reproduite deux fois, en 1989 (3 jeunes ont survécu à l'hibernation) et en 1991 (4), alors que la famille B s'est reproduite 3 fois : en 1989 (1 jeune), 1990 (2) et en 1991 (4). En 1992, aucune des familles ne s'est repro-duite. En 1990, la femelle résidente de B a mis bas, tar-divement, 2 jeunes qui n'ont été observés qu'en août. En 1990, aucune mortalité estivale et hivernale des jeunes n'a été observée. En 1991, 50% des jeunes ont disparu au cours de l'été, les autres ont survécu à l'hibernation.
Activités mensuelles
Les marmottes ont été actives, ces années, 6 mois (début avril à début octobre), et le temps passé hors des terriers a fortement varié d'un mois à l'autre (fig. 1 et 5). Cette variation, cependant, ne dépend pas simplement des moments de sortie et d'entrée journaliers dans les terriers (fig. 1). Une analyse plus détaillée est nécessaire pour mettre en évidence les facteurs responsables de la distribution journalière et mensuelle des activités.
Adultes et un-ans
Comportements actifs/non actifs et temps passé hors des terriers (fig. 1) : En début de saison, les comportements actifs prédominent, puis la tendance s'inverse à partir de juin. Une forte corrélation négative existe entre le temps passé hors du terrier et le pour-centage de pluviosité (rs=-0,829 ; a=0,5).
Comportements non-interactifs :
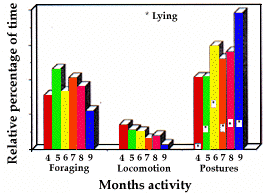
Affouragement (fig. 2). En avril, la durée d'alimentation est relativement faible (31%). Au cours des mois suivants, elle augmente fortement (46%), pour décroître ensuite, et devenir minimum en septembre (22%).
Locomotion (fig. 2). Les pourcentages du temps con-sacré aux déplacements maximum en avril, mai, et juin, respectivement 14%, 11% et 10%, diminuent les mois suivants pour devenir minimum en septembre (3%).
Postures (fig. 2). Le temps passé en position assise ou allongée, minimum en avril et en mai (41%), augmente ensuite jusqu'en septembre où presque 80% du temps leur est consacré. Le pourcentage de temps en position couchée par rapport au temps total de repos est maxi-mum en juin (44%, fig. 2), quand la température journa-lière moyenne (Ta) est de 13deg. C. Au contraire, il est minimum (2,4%) en avril (Ta=1deg. C). En juillet et août (mois les plus chauds, Ta=16 et 18deg. C), le pourcentage moyen de temps passé en position allongée est de 27%.
Récolte et transport d'herbe. Les fréquences (Nombre d'observations/[heures*individu]*100) les plus élevées ont été notées en avril (f=3,6) et en septembre (f=2,8) avec un pic plus faible en juillet (f=1,6). Les autres mois les fréquences sont très faibles, entre f=0 et f=0,8.
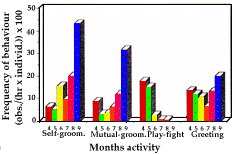
Auto-toilettage (fig. 3). Un maximum marqué apparaît en septembre (f = 43,2) et un minimum en mai (f = 5,2).
Comportements interactifs :
Toilettage mutuel (fig. 3). Il est important surtout en septembre (f=31,6). En avril, il est légèrement plus fré-quent (f=8,8) que les mois suivants.
Interactions de jeu (fig. 3). Très fréquentes en avril (f=17,6) et en mai (15,2), ces interactions décroissent fortement jusqu'en septembre où elles ne sont pas obser-vées. Elles sont en corrélation étroite avec les compor-tements actifs (rs=0,771).
Salutations (fig. 3). Leurs fréquences minimum en juillet, sont relativement élevées tous les autres mois, spécialement en septembre (f=20,0).
Les fréquences des comportements sociaux et de mar-quages, observées en juin au cours des deux années, sont toujours supérieures en 1990. Seules les fréquences mensuelles d'auto-toilettage et de toilettage mutuel sont en corrélation étroites (rs=0,829 ; a=0,5). La corrélation entre les fréquences d'auto-toilettage et le temps passé en posture de repos est élevée (rs=0,943 ; a=0,01), alors que l'hétérotoilettage et les salutations présentent des corrélations modérées avec les postures de repos (respectivement rs=0,657 et rs=0,514).
Interactions agonistiques. Cinq interactions de ce type, impliquant douze animaux, ont été enregistrées. Deux ont eut lieu en mai lorsque des fréquences élevées de marquage ont été observées. Les autres ont eut lieu en avril, juin et août, et à l'exception de cette dernière, elles ont toujours été accompagnées par certaines formes de marquages de la part de l'animal qui a initié le combat.
Comportements de communication
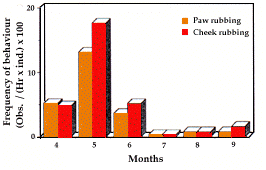
Communication olfactive (fig. 4). Des marquages par frottement des pattes et des joues ont été observés. Ils sont en forte corrélation (rs=0,943 ; a=0,01). Ils présen-tent tous deux des fréquences élevées en début de saison active, avec un pic remarquable en mai. Une forte corrélation a été mise en évidence entre les fréquences de marquage et celles de locomotion (rs=0,771).
Jeunes
Comportements actif/non actif et temps passé hors des terriers (fig. 5) : Ce dernier s'accroît graduel-lement de juin (7,4%) à septembre (31,6%). De même, les comportements actifs et non-actifs augmentent, sauf en septembre. Aucune relation n'a été mise en évidence entre pluviosité et temps passé hors des terriers.
Comportements non interactifs
Affouragement (fig. 6). Il augmente fortement de juin (8%) à juillet (18%), et même plus en août (32%), pour décroître en septembre (12%).
Locomotion (fig. 6). La durée dédiée à la locomotion décroît graduellement de juin (45%) à septembre (12%).
Postures (fig. 6). Comme chez les adultes et les un-ans, les postures de repos atteignent un pic en septembre (66%). La posture allongée est rare chez les jeunes.
Auto-toilettage (fig. 7). En juin et en juillet, les jeunes prennent très rarement soin de leur fourrure (f=2,0). Plus tard, l'auto-toilettage augmente jusqu'à une fré-quence de 10,4 en août et de 12,8 de septembre.
Comportements interactifs
Toilettage mutuel (fig. 7). Ce comportement suit une tendance similaire à celle de l'auto-toilettage.
Interactions de jeu (fig. 7) De fortes fréquences ont été enregistrées en juillet (f = 6,8). Elles sont supérieures chez les jeunes à celles des adultes/un-ans.
Salutations (fig. 7). Elles augmentent tout au long de l'été, avec un minimum en juin (f=1,6), et un maximum en septembre (f=17,2).
L'un des résultats les plus intéressants est l'alter-nance reproductive, observée aussi par d'autres auteurs (Barash 1976, Arnold 1990, Lenti Boero 1991). Il peut s'expliquer en considérant que la femelle résidente ne se reproduit que si sa condition physique, après l'hiberna-tion, lui permet de supporter l'investissement énergé-tique requis par les soins parentaux (Barash 1989). Notre étude permet de préciser cette hypothèse, les femelles se reproduisent seulement lorsque le nombre de jeunes mis bas les années précédentes est de zéro à deux (1989 et 90 pour la famille B ; 1990 pour la famille A). Quand ce nombre est supérieur à 3 (1991 dans la famille B ; 1990 et 92 dans la famille A), les femelles ont une faible probabilité de se reproduire l'année suivante.
Activités mensuelles
Comportements actifs / non actifs et temps passé hors du terrier. Le temps passé hors du terrier par les adultes et les un-ans est fortement influencé par la pluie qui les force à rester dans les terriers. Compte tenu des comportements actifs et non actifs chez les adultes et les un-ans, les mois peuvent être groupés comme suit : 1) les mois actifs : avril et mai ; 2) les mois de faible inactivité : juin, juillet et août ; 3) le mois de forte inactivité : septembre. On doit tenir compte lorsque l'on considère le faible pourcentage de temps passé par les jeunes en juin hors des terriers, que dans cette période, ceux-ci ne s'éloignent guère du terrier (situé sur les déblais) où ils sont difficiles à détecter.
Comportements non interactifs
Affouragement. Sa faible durée en avril est due en par-tie au recouvrement des aires d'alimentation par la neige et à l'importance des activités sociales et d'exploration des marmottes. En mai, ces aires sont presque libres de neige et les animaux s'alimentent près de 50% de leur temps. Ce pic peut s'expliquer par la nécessité pour les marmottes de satisfaire leurs besoins nutritionnels pour se remettre de l'hibernation. En septembre, la faible durée d'alimentation peut être due au fait que la végéta-tion devient sèche, et que les marmottes commencent à ajuster leur métabolisme à des rythmes plus lents.
Locomotion. En avril, malgré le temps passé à se dépla-cer, les marmottes s'écartent peu du coeur du domaine vital. Il est alors possible de formuler quelques hypo-thèses : * La présence de neige oblige les animaux à consacrer plus de temps pour atteindre les aires où la végétation est disponible. * La durée des déplacements est due à la surveillance intensive des frontières du terri-toire (Lenti-Boero 1992). En juin, au contraire, et même plus encore en mai, les déplacements entraînent les adultes et les un-ans dans des aires distantes du coeur du domaine vital. Ces déplacements pourraient être princi-palement le fait des subordonnés explorant les alentours à la recherche d'un territoire où s'installer. Les déplace-ments des jeunes semblent orientés à l'exploration des alentours, quoique parfois il soit difficile de distinguer l'affouragement des activités exploratoires.
Postures. Chez les adultes/un-ans, le temps consacré aux postures de repos augmente au cours de la saison, suivant la tendance biologique de l'été (nombreuses acti-vités au printemps et beaucoup de repos en septembre avant l'hibernation). Les jeunes consacrent en moyenne moins de temps au repos que les autres individus.
Au cours de certaines périodes, les adultes/un-ans se reposent plutôt en position allongée qu'en position assise. Au cours des trois premiers mois de la saison active, le pourcentage de temps consacré à la position allongée semble être influencé par l'élévation de la tem-pérature. Ce résultat renforce l'hypothèse de la fonction de réduction de la température corporelle lors de stress thermiques par la position allongée (Türk & Arnold 1988). Au cours des mois les plus chauds, juillet et août, on s'attendrait à noter une augmentation du pourcentage de cette position alors qu'en fait il décroît. Ceci est pro-bablement dû au fait que les marmottes, lors des heures les plus chaudes de la journée de ces deux mois, entrent dans leur terrier pour se rafraîchir plutôt que de s'allon-ger ou de s'asseoir (Perrin, Le Guelte & Le Berre 1992).
Récolte et transport de l'herbe. Les fortes fréquences observées en début et en fin d'été, reflètent probable-ment le besoin de renouveler la litière de la chambre d'hibernation. Le pic plus faible de juillet pourrait être relié à la récolte d'herbe par les femelles reproductrices.
Auto-toilettage. Fortement lié aux périodes de repos, le pic de septembre, pour les deux classes d'âge, s'expli-querait par l'augmentation de la probabilité des soins de la fourrure lorsque les marmottes passent l'essentiel de leur temps assises ou allongées, et aussi par la destruction des parasites avant l'hibernation. Certains parasites transmettent des maladies (Sabatier 1989), leur destruction accroîtrait la probabilité de survie hivernale lorsque les défenses immunitaires sont réduites.
Comportements interactifs
Toilettage mutuel et auto-toilettage sont en corrélation élevée. Il est donc raisonnable de supposer que ces toi-lettages répondent au même besoin d'éliminer les parasi-tes avant l'hibernation. Les fréquences d'auto-toilettages sont toujours supérieures à celles du toilettage mutuel, sauf en avril. Ces données d'avril pourraient être le résultat du besoin de renforcer les liens sociaux plutôt que de détruire les parasites (auquel cas, un pic compa-rable pour l'auto-toilettage devrait être observé).
Interactions de jeu. Elles sont intenses chez les adultes et les un-ans en avril et en mai, confirmant les observa-tions précédentes (Zelenka 1965, Barash 1976, Sala et al. 1992). Elles pourraient être liées à l'intense activité sociale du début de la saison d'activité. Ce comporte-ment est probablement le meilleur indice pour évaluer les niveaux de l'agitation sociale chez les adultes/un-ans, car c'est le seul comportement social qui semble prendre ses motivations dans la vie sociale. Toilettage mutuel et salutations ont, en plus de leur fonction sociale, une fonction pratique (destruction des parasites et reconnaissance individuelle) qui rend plus difficile la compréhension de leur rôle pendant l'été.
Salutations. Leur importance en septembre s'explique-rait par le fait que les marmottes restant longtemps au repos près des terriers ont plus de chances de se rencon-trer que dans les aires d'alimentation, où elles sont plus dispersées. En début de saison, cependant, postures de repos et salutations ne sont pas en corrélation, suggérant que ces dernières auraient une ou plusieurs motivations additionnelles. Les salutations d'avril et mai sont proba-blement liées à l'activité sociale intense de ces mois. La forte valeur de juin, plus intriguante, peut être reliée au saut de deux mois du cycle reproductif observé en 1990 dans la famille B, ce que confirment les fréquences des autres comportements sociaux et de marquage. Chez les jeunes, les salutations sont rares en juin. Lors des pre-miers jours d'émergence, les contacts avec le reste de la famille ne sont pas établis, et les jeunes restent entre eux ou avec leur mère. Avec l'avancement de la saison, ils deviennent de plus en plus confiants, quelquefois même importuns, et donc leur probabilité de salutations aug-mente jusqu'au maximum de septembre.
Interactions agonistiques. Fortement associées aux marquages, elles servent à la défense du territoire. Elles s'expriment surtout en période de reproduction, quand le mâle résident s'inquiète du cocuage et dans les périodes de forte dispersion (Arnold 1992), quand le territoire doit être défendu contre les individus errants.
Comportements de communication
Communication olfactive. Déplacements et marquages varient de pair, suggérant que ces derniers pourraient être reliées au besoin de défense du territoire. Pour les mâles adultes, l'importance du marquage d'avril, mai, et juin serait liée aux forts taux de testostérone propres à ces mois (Saboureau 1992). Il reste à expliquer pour-quoi les fréquences sont si élevées en mai. Nous devons noter que pendant ce mois, les marmottes errent sur de grandes distances. Donc, les fréquences de marquages peuvent être élevées du fait de la forte probabilité de rencontrer des étrangers errants et, donc, du besoin important de signaler la présence d'un territoire.
Les causes de ces tendances, discutées en détail pour chaque comportement, peuvent se diviser en causes externes et internes. Les facteurs météorolo-giques, les parasites, et la dessication de la végétation sont des facteurs externes qui influencent le cycle estival. Les facteurs internes incluent la reproduction, la dispersion, l'entretien, l'accumulation des réserves de graisses et le besoin de relations sociales.
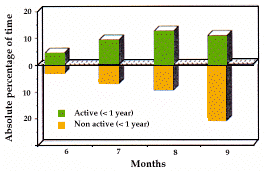
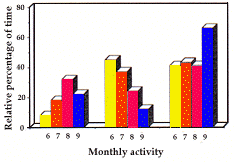
| Figure 1. Durée mensuelle passée hors terrier (activité et non-activité), par les adultes et les un-ans. Les moments de sortie matinale et d'entrée vespérale dans les terriers sont indiqués. / Monthly amount of time spent out of burrow (active and non-active behaviours) by adults and yearlings. Time of morning exit from burrows and evening re-entry are also plotted. | Figure 2. Pourcentages mensuels du temps des comportements d'états par rapport au temps total passé hors terrier. Adultes et un-ans seulement. Monthly relative percentage of time of long lasting behaviours (states) out of the total time spent out of burrow. Adults and yearlings only. |
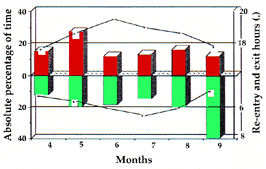 |  |
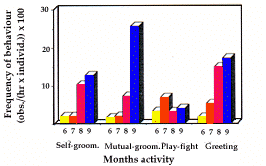
| Figure 3. Fréquences mensuelles des comportements de faible durée (événements) des mâles et un-ans. Monthly short lasting behaviours (events) frequencies of adults and yearlings. | Figure 4. Fréquences mensuelles des marquages.
Monthly frequencies of marking behaviours |
 |  |
| Figure 5. Temps passé hors des terriers par les jeunes chaque mois. Monthly amount of time spent out of burrows by juveniles. | Figure 6. Pourcentages relatifs mensuels de temps passé en
comportements de longue durée (états) chez les jeunes. Monthly
relative percentage of time spent in long lasting behaviours (states) in
juveniles. |
 |  |
Sampling methods. The population, allochtho-nous, was composed of two contiguous families (A and B). The maximum number of recorded animals was 18. Data collection lasted two seasons (1990-91), from March to October, plus two months in 1992 (568 obser-vation hours, 51 days). Field methods consisted mainly in observing the animals with binoculars (7x50 and 8x35) and a monocle (30x100), by a least two observers at a time. Scan and focal sampling methods have been employed (Altmann 1974). Behaviours have been recor-ded according to an ethogram kindly lent by Dr. Lenti-Boero (unpublished paper). Animals have not been marked. Data have been grouped into states (foraging, postures, locomotion, gathering and carrying grass), and events (greetings, play-fighting, self and mutual grooming, paw and cheek rubbing, agonistic inter-actions), according to Lehner (1979). Active behaviours (foraging, locomotion, grass gathering and carrying, and play-fighting) have been distinguished from non active (sitting and lying). Not visible animals have been considered to be in burrows.
Data analysis. Behavioural and meteorological data collected in the field have been coded and entered in a data base. A small part of the data analysis has been carried out by means of the statistical package SPSS, whereas most of the analysis required the elaboration of about 10 original computer programs in FORTRAN 77. Spearman's Coefficient (rs) has been used for correlation.
Out of a period of four years, family A has been observed to reproduce twice, in 1989 (3 offspring sur-vived hibernation) and 1991 (4), while family B repro-duced three times: in 1989 (1), 1990 (2), and 1991 (4). In 1992 neither family reproduced. In 1990 the resident female in family B reproduced very late in the season, delivering two infants that have been observed only in August. In 1990 neither summer nor winter mortality has been recorded in young. In 1991 50% of the off-springs disappeared during summer, but the remaining ones survived hibernation.
Monthly Activities
In both years, marmots have been active for approximately 6 months from early April till early Octo-ber and the amount of time they spent out of burrows varied considerably from month to month (fig. 1 and 5). This variation, however, did not depend simply upon the time of morning exit from burrows and of evening re-entry (fig. 1). It is therefore extremely interesting to pro-ceed with a more detailed analysis in order to under-stand the factors responsible for the monthly and daily distribution of activities.
Adults and yearlings
Active/Non Active Behaviours and Time Spent Out of Den (Fig. 1): Early in the season, active beha-viours prevailed over non active, whereas from June on the trend was reversed. A high negative correlation has been found between the time spent out of the den and the percentage of rainfall (rs=-0.829, a=0.5).
Non Interactive Behaviours
- Foraging (fig. 2). In April, the amount of time spent foraging was relatively small (31%) and increased the following month remarkably (46%), to decrease later on, reaching its minimum in September (22%).
- Locomotion (fig. 2). April, May and June showed the highest percentages of time dedicated to displacements, respectively 14%, 11%, and 10%. Later locomotion reach a minimum in September (3%).
- Postures (fig. 2). The amount of time spent in postures, minimum in April (41%) and May (41%), increased until September, when almost 80% of the time was spent in postures. The percentage of time spent lying out of the total time spent in postures showed a peak in June (44%-fig. 2), when the mean daily air temperature (Ta) was 13deg.C and on the contrary was minimum in April (2.4%) when Ta=1deg.C. In July and August (the warmest months, with respectively Ta=16deg.C and Ta=18deg.C), the average percentage of time spent lying was 27%.
- Gathering and carrying grass. The highest frequencies have been recorded in April (f=3.6 (ndeg. observations / (hour*individual))*100) and September (f=2.8), with a minor peak in July (f=1.6). Other months showed very low frequencies ranging from f=0.0 to f=0.8.
- Self-Grooming (fig. 3). It showed a sharp maximum in September (f=43.2), and a minimum in May (f=5.2).
Interactive Behaviours
- Mutual Grooming (fig. 3) was performed mainly in September (f=31.6) and slightly more in April (f=8.8) than in the following months.
- Play-fighting (fig. 3). Very high frequencies in adults and yearlings have been exhibited in April (f=17.6) and May (f=15.2). The following months, frequencies de-creased considerably till September when no play-fighting has been recorded. Play-fighting correlated strongly with the trend of active behaviours (rs=0.771).
- Greetings frequencies (fig. 3) showed a minimum in July (f=6.4), with relative high values in all the other months and especially in September (f=20.0).
-The frequencies of social and marking behaviours observed in June in the two different years were always higher in 1990. Only self-grooming and mutual groom-ing turned out to be strongly correlated (rs=0.829, a=0.5). Frequencies of self-grooming were highly cor-related with the percentage of time spent in postures (rs=0.943, a=0.01), while mutual grooming and greet-ings were only moderately correlated with postures (respectively rs=0.657, and rs=0.514).
- Agonistic interactions have been recorded five times, involving twelve animals. Two of them took place in May, when high frequencies of marking behaviours have also been observed. Others observed in April, June and August, were always accompanied by some kind of marking activity performed by the individual that started the fighting, except in August.
Communicative Behaviours
- Olfactive communication (fig. 4). The marking pat-terns of cheek and paw rubbing have been observed and were strongly correlated (rs=0.943, a=0.01). Both showed very high frequencies in the first part of the active season, with a remarkable peak in May. A consi-derable correlation has been found between marking frequencies and locomotion (rs=0.771).
Infants
Active/Non Active Behaviours and Time Spent Out of Den (fig. 5): The time which the juveniles spent out of the den increased gradually from June (7.4%) to September (31.6%-fig. 5). According to this trend there was an increase both in active and non active behaviours, except for September. No relation has been found between rainfall and time spent out of burrows.
Non Interactive Behaviours
Foraging (fig. 6) increased remarkably from June (8%) to July (18%), and even more in August (32%), to decrease in September (22%).
Locomotion time (fig. 6) decreased gradually from June (45%) to September (12%).
Postures (fig. 6). In infants, as well as in adults and yearlings, postures reached a peak in September (66%). Lying has been observed very seldom in infants.
Self-Grooming (fig. 7). In June and July young marmots took care very seldom of their fur (f=2.0). Later on self-grooming increased to a frequency of 10.4 in August and of 12.8 in September.
Interactive Behaviours
Mutual grooming (fig. 7) followed a trend which is similar to that of self-grooming.
Play-fighting (fig. 7). High frequencies of play-fight have been recorded in July (f=6.8). During the active season, young marmots showed play frequencies higher than those of adults and yearlings.
Greetings (fig. 7) showed a growing pattern through-out the summer, with a minimum in June (f=1.6), and a maximum in September (f=17.2).
One of the most interesting results is the skipping of reproductive seasons which has been observed also by other authors in alpine marmot (cfr. Barash 1976, Arnold 1990; Lenti Boero 1991). This fact may be ex-plained by considering that the resident female repro-duces only if her physical conditions, after hibernation, are sufficient to support the energy investment required by parental cares (Barash 1989). From our study it is possible to put forward a slight quantitative improve-ment of this hypothesis: females reproduce only when the number of infants delivered in the previous year ranges from zero to two ('89 and '90 in family B; '90 in family A). When the number of infants is greater than 3 ('91 in family B; '90 and '92 in family A) females are not likely to reproduce in the following year.
Monthly Activities
Active/Non Active Behaviours and Time Spent Out of Den . The amount of time spent out of the den by adults and yearlings was strongly influenced by rain, which forced the animals to spend more time in bur-rows. With respect to active and non active behaviours in adults and yearlings, months can be grouped as follows: 1) Active months: April and May; 2) Slightly inactive months: June, July, and August.; 3) Strongly inactive months: September. Care must be taken when considering the low percentage of time spent out of the den by infants in June, since in this first period infants did not move far away from the den (which is on detritus) and were therefore difficult to detect.
Non Interactive Behaviours
Foraging. The low amount of time spent foraging re-corded in April was in part due to the fact that foraging areas were still covered by snow and that marmots were very busy with social and explorative activities. In May feeding areas were almost free from snow, and the ani-mals spent almost 50% of their time foraging. The peak observed in May might be explained with the necessity of marmots to satisfy nutritional needs in order to reco-ver completely from hibernation. In September, the low amount of time spent feeding might be due to the fact that vegetation was becoming dry, and that marmots started to set their metabolism on more slow rhythms.
Locomotion. In April, in spite of the relatively great amount of time dedicated to locomotion, marmots did not wander far away from the core of the home-range. This makes possible to formulate some hypothesis: * Snow forced the animals to employ more time to reach areas with available vegetation. * The time employed in displacements was due to intensive patrolling of the territorial boundaries (cfr. Lenti-Boero 1992). In June, on the contrary, and even more in May, displacements led adults and yearlings to reach very distant areas from the core of the home-range. These displacements may be due mainly to subordinates exploring the sur-roundings in search for a territory where to become residents. In infants, displacements seemed to be aimed at exploring the surroundings, although it was some-times very difficult to distinguish between foraging and explorative activities in young.
Postures. In adults and yearlings the time spent in postures increased as the season proceeded following the evolution of the biological summer trend, which was characterised by a lot of activity in spring and by a lot of resting in September before winter sleep. Young spent on average less time in postures than adults and yearlings. During certain periods, adults and yearlings spent a lot of their resting time lying rather than sitting. During the first three months of the active season the percentage of time spent lying seemed to be influenced by the increasing temperature. This result agrees with the hypothesis made by Türk & Arnold (1988) about the function of lying, as a mean of lowering the body tempe-rature when in heat stress. In July and August, the warmest months, we expected an increase of lying percentage, instead it decreased. This was probably due to the fact that during the warmest hours of the day, of both July and August, marmots rather re-entered burrows in order to cool down, than stay out lying or sitting (cfr. Perrin, Le Guelte, & Le Berre 1992).
Gathering and carrying grass. The high frequencies recorded at the beginning and at the end of the summer season, probably reflect the need for renewing the litter of the hibernation room. A minor peak in July may be attributed to the grass collected by the reproductive female for her offspring.
Self-Grooming was strongly related with periods of rest. In particular the peak in September, characteristic of both age classes, may be explained by the fact that marmots spent most of their time sitting or lying, and therefore had many chances to take care of their fur. This peak may also be attributed to the fact that mar-mots try to get rid of the parasites before winter sleep. Some parasites carry diseases (for example Sabatier 1989) and therefore getting rid of them might increase the probability of survival during hibernation, when a lowering of the immune defences is to be expected.
Interactive Behaviours
Mutual Grooming. Frequencies of mutual and of self-grooming were remarkably correlated. It is therefore reasonable to suppose that mutual grooming was brought about by the same need of eliminating para-sites, especially before winter sleep. Except for April, frequencies of self-grooming were always higher than those of mutual grooming. April's datum should be attributed to the need of consolidating social bonds, rather than to the need of getting rid of the parasites (we would otherwise expect a comparable peak in self-grooming, which has not been observed).
Play-fighting. Intense play-fighting has been observed in adults and yearlings during April and May, in agree-ment with observations by other authors (Zelenka 1965, Barash 1976, Sala et al. 1992). Such a frequency may be explained by considering the intense social activity that took place at the beginning of the active season. Play-fighting is probably the best index to evaluate levels of social agitation in adults and yearlings, because it is the only social behaviour that seems to find all its motivations in social life. Mutual grooming and greetings have, besides a social function, also a practical function (getting rid of parasites, and individual recognition, respectively), which makes it more difficult to under-stand their role during summer.
Greetings highly frequent in September may be ex-plained by considering that marmots spent most of their time sitting and lying near burrows, and therefore had more chances to meet each other than in the feeding areas, where they were more dispersed. In the early part of the summer season, however, greetings were not correlated with resting, suggesting that there should be one or more additional motivations. In April and May, a lot of greeting was probably due to the intense social activity characterising these months. The high value found in June is more intriguing, and might be related to the two months' shifting of the reproductive cycle re-corded in 1990 in family B. This tendency is confirmed when considering the frequencies of the other social behaviours and of marking behaviours. In infants, greetings have been observed very seldom in June. In their first days of open air life, contacts with the rest of the family were not completely established, and offspring spent most of their time among them or with their mother. As the season proceeded infants became more and more confident, sometimes even intrusive, and therefore the chance for greetings increased until September, when they reached a peak.
Agonistic interactions were strongly associated with marking behaviours. They are aimed at defending the territory and took place mainly in the periods of repro-duction, when the resident male is afraid of cuckoldry, and in the period of highest dispersion (cfr. Arnold 1992), when the territory must be defended from wandering individuals.
Communicative Behaviours
Olfactive communication. Elevated values of marking behaviours were associated with high values of displa-cements, and this suggests that markings may be related to the need of defending the territory. In the adult male, the high frequencies of marking observed in April, May, and June could be related with the high rate of testo-sterone typical of these months (Saboureau 1992). It must still be explained why frequencies were so high in May, when compared with April and June. To this pur-pose it must be pointed out that in May marmots wandered great distances. Markings may have been so frequent because there was a higher probability of meeting wandering strangers, and therefore a higher need of signalling the presence of a defended territory.
The causes of these trends, discussed in detail for every single behaviour, can be divided into external and internal. Meteorological parameters (temperature, rain and snow), parasites, and vegetation dryness are among the external factors influencing summer cycle. Internal factors include reproduction, dispersion, maintenance, fat accumulation, and the need for social relations.
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |