![]()
Cinq groupes sociaux de marmottes identifiées individuellement ont
été observés dans le Parc National du Grand Paradis,
Italie, de l'été 1985 à 1988. La taille du domaine vital
varie de 12.500 m2 à 28.125 m2 (m.=18.125
m2). Le coeur du domaine, où 10% des positions ont
été observées, a en moyenne 1.818 m2. Dans
chaque domaine, un seul terrier d'hibernation est utilisé au cours des
années par différents groupes sociaux, le nombre moyen de
terriers utilisés pendant l'été a été de
2,81 (1 à 6). Les sites de surveillance hors terriers varient de 2
à 13% (m. 5,64%) et se trouvent à la périphérie des
domaines. Aucune relation n'a été mise en évidence entre
le nombre d'animaux dans un groupe et la taille du domaine ou entre la taille
du domaine et le nombre de sites d'observation. La plasticité
comportementale de l'utilisation des ressources spatiales et les facteurs
sociaux et écologiques influençant les dimensions du domaine
vital et du nombre des autres ressources sont discutés.
Mots-clés : Marmotte alpine, Marmota marmota, Alpes occidentales, Parc national du Grand Paradis, domaine vital, plasticité comportementale.
Five social groups of individually identified marmots were observed at Gran
Paradiso National Park, Italy from Summer 1985 to 1988. Home range size varied
from 12.500 sm to 28.125 sm (mean = 18.977). Core areas, where at least 10% of
fixes were found had a mean size of 1.818 sm. In each home range only one
hibernating burrow utilized through years by different social groups was found,
the mean number of burrows used during summer was 2.81 (range 1 - 6). Spotting
points, that is rocks or logs without burrows varied from 2 to 13 (mean 5.64%)
and were located at the periphery of the home ranges. No relationship was found
between number of animals in a social group and home range sizes or between
home range sizes and number of observation places. Behavioural plasticity in
use of spatial resources and the social and ecological factors influencing
dimensions of home ranges and number of other resources is discussed.
Key-words: Alpine marmot, Marmota marmota, Western Alps, Gran Paradiso National Park, Italy, home range, behavioural plasticity.

La survie des marmottes alpines dépend des res-sources hivernales, surtout d'un hibernaculum efficace (Arnold 1990, 1992, 1993), et de ressources estivales telles que le fourrage et les terriers-abris jour et nuit. Avec d'autres facteurs du milieu comme les points de surveillance (Bopp 1954, 1956, 1958, Zelenka 1965, Lenti Boero 1985, 1995, Perrin et al. 1993) et les pierres et bouts de bois utilisés comme site de marquage (Lenti Boero 1993, 1995) ces ressources sont réparties dans les domaines vitaux, qui deviennent des territoires au cours du printemps (Zelenka 1965) ou tout au long de l'année et qui sont utilisées par des individus différents de chaque groupe social. Les dimensions de ces domaines varient suivant la distribution de la population : ceux des groupes familiaux isolés sont plus grands que ceux des groupes en colonie (Bopp 1954, 1956, 1958, Zelenka 1965, Lenti Boero 1985, 1995, Mann & Janeau 1988, mais voir Perrin et al. 1993) et probablement aussi en fonction de facteurs écologiques (Bopp 1954, 1956, Perrin et al. 1993). Les données concernant les domaines vitaux, l'utilisation des terriers et les points de surveillance sont présentées pour une colonie vivant dans un milieu extrême (Lenti Boero 1994) au Parc National du Grand Paradis (PNGP), Italie.
Site d'étude et recueil des données : Les données ont été récoltées de 1985 à 1988 de juillet à début septembre, dans un site situé à 2300 m, au-dessus de la limite des forêts dans une petite vallée latérale à la vallée principale de Cogne (PNGP), vallée d'Aoste, Italie (Lenti 1985, Lenti Boero 1991, 1992). La surface totale du site a été subdivisée en une grille de 114 carrés de 25 m de côté. Chaque carré est coté par une lettre et un nombre. La matrice a été surimposée à une photogra-phie du site d'étude (Barash 1973), d'une surface de 7,6 ha. Au cours de l'étude, elle a été occupée par trois domaines vitaux distincts : FamEl, FamGia et FamGenz (Lenti Boero 1991). En 1988, deux groupes sociaux ont été remplacés par deux couples d'immigrants extérieurs à la colonie. Aussi, 5 groupes sociaux différents ont été observés pendant toute l'étude (Lenti Boero & Boero 1989, 1991, 1994).
Sujets et recueil des données : 22 marmottes adultes et subadultes et 13 marmottons ont été capturés et marqués pour une reconnaissance individuelle, et 12 adultes et 5 marmottons non marqués ont été suivis. L'observateur a suivi, d'un lieu peu visible, au téléscope 30x60 et aux jumelles 10x40 les animaux. Les données sur l'utilisation de l'espace de chaque groupe familial ont été collectées au cours de 1302 échantillons par balayage visuel, de 10 minutes chacun. La durée totale d'observation a été de 217 heures (Lenti Boero 1991). De plus, des données ont été collectées par échan-tillonnage focalisé sur un animal (Altmann 1974) au cours de 221 heures. Les dimensions des domaines vitaux ont été estimées directement à partir des fré-quences des positions dans chaque carré, recueillies pour chaque groupe social (Macdonald et al. 1980). Les données ont été analysées avec Systat.
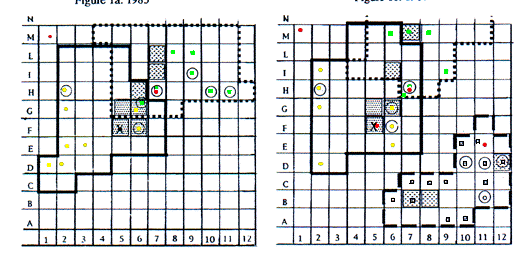
Les caractéristiques des domaines vitaux, des ter-riers inclus, des points de vue sont présentées fig. 1a, b, c, d. La forme des domaines a varié au cours de l'année pour un même groupe social comme pour les nouveaux arrivants, mais, le coeur du domaine, où sont comptées plus de 10% des positions, est resté constant ou s'est peu déplacé au cours des années. Cette zone comprend le système principal de terrier (SPT, Perrin et al. 1993) ou les zones d'affouragement très fréquentées.
Les dimensions totales du domaine vital varient au cours des années (tab. 1), suite à une réduction reliée à la substitution d'un nouveau couple et une diminution des animaux dans le groupe social, mais sans relation significative (test de l'effet du nombre d'individus sur les dimensions du domaine vital : F = 2,11, P = 0,18, ns et F = 2,72, P = 0,13, ns, F = 2,36, P = 0,16, ns, respectivement pour le domaine vital entier, pour le coeur et la racine carré du total). Cela suggère l'exis-tence d'autres facteurs qui pourraient influer sur les di-mensions du domaine vital, probablement des facteurs sociaux liés au comportement des animaux voisins, et/ou des facteurs écologiques en liaison avec des carac-téristiques alimentaires.
Un seul terrier d'hibernation, ressource perma-nente, a été trouvé dans chaque domaine vital. Dans le domaine de FamEl, de 1985 à 1987, ce terrier a été fer-mé avec de la terre et des pierres et ouvert seulement début septembre. J'ai creusé l'entrée en juillet 1986, mais les mâles l'ont rapidement rebouché. Dans les autres domaines, les terriers d'hibernation étaient uti-lisés aussi pendant l'été.
Tous les animaux d'un groupe utilisent librement le SPT et les points de vue situés près des terriers ou sur des rochers proches ou aux frontières des domaines, quelquefois à 100 m ou plus du SPT. Différentes struc-tures d'observation ont été utilisées au cours des années mais un seul SPT par domaine vital (25% de tous les terriers), et quelques points de vue (9,5% de l'ensemble) l'ont été de façon permanente. Ceux-ci, avec le terrier d'hibernation, semblent être des ressources normales du domaine vital, et ont été utilisées successivement par les différents groupes sociaux qui les ont occupées. La rela-tion entre étendue et nombre de sites d'observation n'est pas significative (F = 0,55, P = 0,078). En fait, la cor-rélation entre ces points d'observation et le danger d'intrusion des animaux voisins pourrait être meilleure.
Les domaines vitaux de marmottes alpines vivant en colonie ont été mesurés (Bopp 1954, 1956, 1958, Zelenka 1965, Mann & Janeau 1988, Perrin et al. 1993), et l'hypothèse d'une relation entre les stratégies d'occu-pation de l'espace, la taille des domaines vitaux et les ressources alimentaires a été avancée par Mann & Janeau (1988). Il est possible de distinguer les canevas d'alimentation suivants : "l'alimentation ordinaire", les animaux se déplacent le long d'un sentier et choisissent sur une surface réduite les plantes à consommer et "l'ali-mentation spécifique", les animaux voient une plante intéressante et se déplacent hors du sentier pour l'attein-dre (Lenti Boero in prep.), ce qui suggère l'existence de plantes ayant un intérêt spécial. Une forte proportion des positions d'animaux broutant hors du coeur du domaine sont des animaux s'alimentant (Lenti Boero in prep.), impliquant que les carrés de la grille hors coeur du domaine ont été explorés à la recherche de nourriture ou de ressources particulières répandues sur le territoire. Frase & Armitage (1989) ont montré que les marmottes sont des herbivores généralistes, mais sans exclure que certains items alimentaires puissent être spécifiques (Armitage comm. pers.), seule une meilleure connais-sance du régime alimentaire permettra de clarifier ce point. Cependant, les activités des marmottes au cours de l'été ne se limitent pas à l'alimentation mais aussi à la défense contre les prédateurs et contre les intrusions de conspécifiques. Les bains de soleil ou la surveillance sur les terriers pourraient avoir cette double fonction. Ce-pendant 39,5% de toutes les positions observées l'ont été sur des sites d'observations éloignés de terriers, dans ce cas le comportement est dirigé vers la surveillance des intrus, et des prédateurs ont été vus très souvent dans le voisinage (Lenti Boero 1991, 1992), d'autant que sur-veiller à partir d'un site visible est certainement un comportement à risque.
En conclusion, cette exploration préliminaire des données suggère qu'une multiplicité de facteurs, tels que le type de nourriture et l'intrusion sociale interagissant probablement avec le nombre d'animaux du groupe pourrait influer sur les dimensions des domaines vitaux et sur la dynamique d'utilisation de l'espace.
| Figure 1a. 1985 | Figure 1b. 1986 |
| Figure 1c. 1987 | Figure 1d. 1988 |
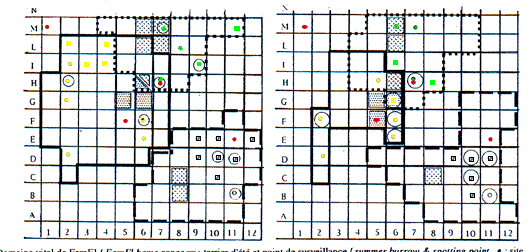
Domaine vital de FamEl / FamEl home range ; terrier d'été et point de surveillance / summer burrow & spotting point ; site de surveillance / spotting point ; sites de surveillance ou de bain de soleil où plus de 10% des observations ont été réalsiées / more than 10% of fixes when spotting or sunning ; sites d'afourragement ou de déplacement où plus de 10% des observations ont été réalsiées / more than 10% of fixes when foraging or walking .
Domaine vital de FamGia / FamGia home range ; terrier d'été et point de surveillance / summer burrow & spotting point ; site de surveillance /spotting point ; sites de surveillance ou de bain de soleil où plus de 10% des observations ont été réalsiées / more than 10% of fixes when spotting or sunning ; sites d'afourragement ou de déplacement où plus de 10% des observations ont été réalsiées / more than 10% of fixes when foraging or walking .
Domaine vital de FamGenz /FamGenz home range ; terrier d'été et point de surveillance / summer burrow & spotting point ; site de surveillance / spotting point ; sites de surveillance ou de bain de soleil où plus de 10% des observations ont été réalsiées / more than 10% of fixes when spotting or sunning ; sites d'afourragement ou de déplacement où plus de 10% des observations ont été réalsiées /more than 10% of fixes when foraging or walking .
Tableau . Données brutes et paramètres des domaines vitaux au PNGP en 1985-1988
Table 1. Row data and parameters of home ranges at PNGP 1985-1988
| Group and year* | N. of animals | Total home range | Home core (> 10% of fixes) | N. of MBS including sp. | N. of sp out of burrows | Total sp. points |
| FamEl 1985 | 9 | 23125 | 2500 | 6 | 3 | 9 |
| FamEl 1986 | 6 | 24375 | 1875 | 5 | 3 | 8 |
| FamEl 1987 | 9 | 28125 | 1875 | 5 | 10 | 15 |
| FamPal 1988 | 3 | 12500 | 1875 | 3 | 2 | 5 |
| FamGia 1985 | 4 | 21250 | 1875 | 1 | 9 | 10 |
| FamSo 1986 | 2 | 16875 | 1875 | 2 | 3 | 5 |
| FamSo 1987 | 2 | 13125 | 3125 | 3 | 4 | 7 |
| FamElid 1988 | 2 | 15625 | 1875 | 3 | 3 | 6 |
| FamGenz 1986 | 10 | 15625 | 1875 | 1 | 13 | 14 |
| FamGenz 1987 | 12 | 18750 | 1250 | 1 | 5 | 6 |
| FamGenz 1988 | 14 | 19375 | 625 | 1 | 7 | 8 |
| range | 12 | 15625 | 2500 | 5 | 11 | |
|---|---|---|---|---|---|---|
| mean | 645.8 | 18977.27 | 1818.18 | 2.81 | 5,64 | |
| SD | 4.46 | 4883.58 | 652.58 | 1.83 | 3.61 | |
| SE | 1.34 | 1472.45 | 196.82 | 0.55 | 1.09 | |
Alpine marmots depend for their survival both on winter resources, mainly efficient hibernating burrows (Arnold 1990, 1992, 1993) and summer resources such as forage and burrows utilized as day and night shelters. Together with other environmental cues such as spotting points (Bopp 1954, 1956, 1958, Zelenka 1965, Lenti Boero 1985, 1995, Perrin et al. 1993) and stone or logs used as marking sites (Lenti Boero 1993, 1995) those resources are distributed within home ranges, which become defended territories during springtime (Zelenka 1965) or throughout the year and are utilized by different individuals within each social group. Dimensions of home ranges vary according to the distribution of the population: isolated family groups utilize wider home ranges than numerous social groups living in colonies (Bopp 1954, 1956, 1958, Zelenka 1965, Lenti Boero 1985 and 1995, Mann & Janeau 1988, but see Perrin et al. 1993) and also probably according to ecological factors (Bopp 1954, 1956, Perrin et al. 1993). In this paper data on dimensions of home ranges and utilization of burrows and spotting points are presented for a colony living in an extreme environment (Lenti Boero 1994) at the Gran Paradiso National Park, Italy.
Study Area and data collection: Data were collected from 1985 to 1988 during the months of July, August and early September. The study area was located at 2300 m, above timberline in a small valley branching from the main Cogne Valley at the Gran Paradiso National Park, Valle d'Aosta, Italy. For further details see Lenti (1985) and Lenti Boero (1991, 1992). In order to determine home ranges space use the entire area was subdivided in a grid of 114 squares with 25 m on each side, with the aid of a tape measure. Each square was denoted with a letter and a number and the whole matrix was superimposed on a photograph of the study area as suggested by Barash (1973). The surface covered by the grid measured 7.6 ha. During the study the area supported three distinct homes ranges: FamEl, FamGia and FamGenz (Lenti Boero 1991). In 1988 two social groups were completely replaced by two pairs immigrating from outside the colony, consequently, a total of five different social groups were observed during the entire study (for further details see Lenti Boero and Boero 1989, 1991, 1994).
Subjects and collection of data: 22 adult and subadult marmots and 13 new-borns were live trapped and marked for individual recognition. In addition 12 unmarked adults and 5 new-borns were observed.
The observer sat in an inconspicuous place with 10X40 binoculars or a 30X60 telescope. Information about space use of individuals belonging to each family group was gathered on selected days by means of 1302 scan samples, lasting 10 min each. The total amount of time spent in scanning was 217 h (Lenti Boero 1991). In addition, observational data were collected by means of focal animal sampling (Altmann 1974) in 221 h of direct observation.
Dimensions of home ranges were estimated di-rectly from the frequency of fixes in each grid gathered for each social group as suggested by Macdonald et al. (1980). Data were analysed with Systat.
Characteristics of home ranges and included burrows and spotting points are presented in fig. 1a,b,c,d. Shapes of home ranges varied across the year for the same social group and for newcomers, however, the core area, where more than 10% of fixes were counted, was permanent or slightly displaced across years. The area was located around the main burrow system (Perrin et al. 1993) or in highly frequented foraging areas.
Total home range dimensions varied across years (tab. 1), with a reduction connected with the substitution of a new pair and a consequent reduction of animals in the social group, but no significant statistical relationship was found (test for the effect of number of animals on home range dimensions: F = 2.11, P = 0.18, ns and F = 2.72, P = 0.13, ns, F = 2.36, P = 0.16, ns, for total home range, core area and square root of total respectively). This suggests the existence of other factors that might influence home range dimensions, possibly social factors related with behaviour of neighbouring animals, and/or ecological factors related with food characteristics.
Hibernating burrows were permanent resources and only one was found in each home range. In FamEl home range, from 1985 to 1987, the hibernating burrow was closed with ground and stones and opened only in early September. I excavated the entrance in July 1986, but males from the group soon closed it again. Hiber-nating burrows in other home ranges were utilized also during the summer.
All animals in a group freely utilized the MBS and the spotting points located at the entrances of bur-rows, or on rocks near them or near the boundaries of home ranges, sometimes 100 m or more away from the MBS. Different observational structures were used across years and only one MBS in each home range (25% of all burrows found), and few spotting points (9.5% of all spotting points found) were permanently utilized during all the study. Those, together with the hibernating burrow, seemed to be the "regular resources" of each home range, and were successively used by the different social groups successively inhabiting them. The Pearson correlation coefficient between range and number of observation spots was not significant (F = 0.55, P = 0.078), in fact, observation spots might be better correlated with the danger of intrusion of neighbouring animals.
Home range measures for colonies of alpine marmots have been reported by other authors (Bopp 1954, 1956, 1958, Zelenka 1965, Mann & Janeau 1988, Perrin et al. 1993), and the hypothesis of a relationship between space occupation strategies, home range sizes and foraging resources has been advanced by Mann & Janeau (1988). Foraging patterns can be distinguished in "ordinary forage", when animals walk along a predefined path and choose in a small area the plants to be eaten, and "specific forage", when animals see at a distance an interesting plant and displace themselves from the path, in order to reach it (Lenti Boero in prep.), this suggests the existence of particular food of specific interest. A great proportion of the fixes of ani-mals grazing outside the core area was of foraging ani-mals (Lenti Boero in prep.), this also indicates that squares of the grid outside the core area are explored in order to gather food or critical dietary resources wide-spread on the territory. Frase & Armitage (1989) show-ed that marmots are generalist herbivores, but this does not preclude that certain food items can be critical (Armitage pers. comm.), only further knowledge on diet will clarify this point. However, marmot summer activi-ties are not only related with foraging but also with defence against predator and conspecific intruders. Sunning or spotting at burrows might have the double function, however 39.75% of all fixes were found on spotting points without burrows, in this case the behaviour is aimed at surveillance against intruders, in fact predators were seen in the area quite frequently (Lenti Boero 1991, 1992), and spotting in a conspicuous place away from a burrow is certainly a behaviour at risk.
In conclusions, this preliminary exploration of spatial data suggests that a multiplicity of factors, namely type of food and social intrusion possibly interacting with number of animals in a group might influence the dimensions of home ranges and the dynamic of space utilization.
ALTMANN J. 1974. Observational study of behaviour: sampling methods.
Behav., 49: 227-265.
ARNOLD W. 1990b. The evolution of marmot sociality: 2. Costs and benefits of
joint hibernation and helping. Behav. Ecol. Sociobiol., 27: 239-246.
ARNOLD W. 1992. Social evolution and obligatory group hibernation in marmots.
In Proc. 1st Int. Symp. Alpine Marmot, Bassano B., Durio P., Gallo Orsi
U., Macchi E., eds., Torino, 41-54.
ARNOLD W. 1993. Social evolution in marmots and the adaptive value of joint
hibernation. Verh. Dtsch. Zool. Ges., 86: 79-93.
BARASH D.P. 1973. The social biology of the Olympic Marmot. Anim. Behav.
Monogr., 3.
BONESI L. 1993. Marmota m. marmota (Linnè 1758): Biologia del
comportamento e utilizzo dello spazio di una popolazione alloctona delle
Prealpi Giulie. Unpubl. thesis. University of Trieste.
BOPP P. 1954. Zur Topographie der Murmeltierterritorien. Rev. Suisse
Zool., 61: 374-380.
BOPP P. 195. Zur Topographie eines Kolonialterritoriums bei murmeltieren.
Rev. Suisse Zool. , 63: 255-261.
BOPP P. 1958. Fluchtdistanzen und territoriales Verhalten beim Murmeltier.
Mitt. Naturforsch. Ges., 26: 1-6.
FRASE B.A. JR. & ARMITAGE K. B. 1989. Yellow-bellied marmots are generalist
herbivores. Ethol. Ecol. & Evol., 1: 353-366.
LENTI D. 1985. Ecologia degli insediamenti e utilizzo delle tane nella marmotta
alpina (Marmota marmota L.). In Att. IV Congr. Naz. Div. Ric. Bas.
Psicol, Ravello, 35-36.
LENTI BOERO D. 1991: Permanenza sui territori, avvicendamento di gruppi
familiari, natalità e sopravvivenza in una colonia di marmotta alpina
(Marmota marmota L.): dati preliminari. In Suppl. Ric. Biol. Selv.
XI,. Bologna, 461-474.
LENTI BOERO D. 1992. Alarm calls in marmots: evidence for semantic
communication. Ethol. Ecol. & Evol, 4: 125-138.
LENTI BOERO D. 1993. La funzione comportamento di marcatura nella marmotta
alpina nella comunicazione olfattiva di possesso del territorio. Ric.
Psicol., 17: 75-90.
LENTI BOERO D. 1994. Survivorship among young Alpine marmots and their
permanence in their natal territory in a high altitude colony. Ibex,
JME, 2: 9-15.
LENTI BOERO D. 1995. Scent Deposition Behaviour in Alpine Marmots (Marmota
marmota L.): its role in territorial defence and social communication.
Ethology, 100: 25-38.
LENTI BOERO D &, BOERO E. 1989. Dati preliminari sulla dispersione e
predazione nella marmotta alpina (Marmota marmota L.): uno studio a
lungo termine. In Att. VIII Congr. Naz. Div. Ric. Bas. Psicol., Padova,
57-59.
MACDONAL D. W., BAL, F. G. & HOUGH N. G. 1980. The evaluation of home range
size and configuration using radio tracking data. In A handbook of
biotelemetry and radiotracking, Jamlaner C. & MacDonald D., eds.,
Pergamon Press, Oxford, 405-424.
MANN C. S. & JANEAU G. 1988. Occupation de l'espace, structure sociale, et
dynamique d'une population de marmottes des Alpes. Gib. Faun. Sauv.,
5: 427-445.
PERRIN C., ALLAINE D. & LE BERRE M. 1993. Socio-spatial organization and
activity distribution of the alpine marmot Marmota marmota: preliminary
results. Ethol., 93: 21-30.
ZELENKA G. 1965. Observations sur l'écologie de la marmotte des Alpes.
La terre et la Vie, 19: 238-256.
A suivre
Forward
![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |