![]()
Les réactions de M. bobac au danger et au signal d'alerte sont
étudiées en fonction de l'âge. Les jeunes (3 mois)
très réactifs émettent le signal d'alerte à la vue
de presque tout objet mobile. Plus intensément sur leurs gardes que les
adultes, ils reprennent plus vite leur activité interrompue. Leur signal
se distingue de celui des adultes par sa fréquence dominante et maximale
et par la durée plus longue de sa composante à haute
fréquence. Les marmottes des 2 groupes d'âge distinguent les
signaux d'un adulte de ceux d'un jeune, ignorent plus souvent les derniers et
restent plus longtemps sur leurs gardes en réponse aux premiers. Le
signal de danger, émis par un jeune, étant souvent peu fiable, la
réaction plus faible à ce signal paraît biologiquement plus
utile, permettant aux marmottes de consacrer plus de temps aux activités
positives, surtout l'alimentation.
Mots-clés : Marmotte des steppes, Marmota bobac, communication acoustique, comportements
dépendant de l'âge, cri d'alarme, structure spectrale.
Age-dependant features M. bobac response to a danger and to the
species alarm call were investigated. Cubs (3 months old) differ by higher
reactivity, giving an alarm call to almost every approaching object. They are
significantly intensely alerted in response to alarm call, but quicker resume
interrupted activity than elder marmots. Their call differs from adults' one by
higher dominant and maximum frequencies and longer duration of the
high-frequency component. Animals of both age groups, discriminate adult and
juvenile signals, more often ignoring the latter and staying longer alerted in
response to the former. Inasmuch as alarm signal given by a juvenile may often
transmit unreliable information, less active response to these calls seems to
be biologically beneficial, permitting marmots to spare more time to the other
positive forms of activity, such as foraging.
Key-words: Steppe marmot, acoustic communication, age-dependent behaviour, alarm call, spectral structure.

Ce travail s'inscrit dans une d'étude à long terme sur la capacité des marmottes, d'une part, à reconnaître leur propre cri de ceux d'espèces voisines ou distantes, d'autre part, à discriminer les situations et la variabilité des cris d'alarme de conspécifiques. Marmota bobac, M. baibacina et M. caudata ont été étudiées (Nikol'skii & Nesterova 1988, 1989, 1990, 1991). Le but de cette étu-de est de mettre en évidence les différences de réponse aux cris d'alarme de conspécifiques en fonction de l'âge, ainsi que d'examiner la sensibilité des marmottes aux critères d'âge de la structure spectrale du cri d'alarme en utilisant le modèle de la repasse sur le terrain.
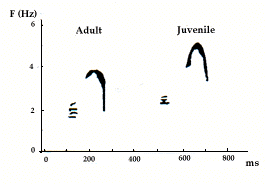
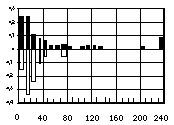
Les réponses des marmottes à des stimulus sono-res artificiels ont été testées à l'aide d'un magnétophone portable émettant des cris d'alarme préenregistrés dans cette population et artificiellement arrangés : 1) ensem-bles de sons de 30 s construit à partir de répétitions du cri naturel d'alarme d'un adulte espacées de 4 s, 2 s et 0,6 s afin de révéler les différences de comportement d'alerte entre juvéniles et adultes ; 2) des cris d'alarme émis par des animaux d'âge différent, pour tester l'in-fluence du facteur âge. Les cris d'alarme de ces mar-mottes forment une séquence de sons séparés (cris) par des intervalles de quelques centaines de millisecondes à plusieurs secondes, chacun comprenant 2 composantes : une basse et une haute fréquence. L'analyse sonagra-phique montre que cette dernière a, chez les juvéniles, une dominante plus forte et une durée plus longue que chez les adultes (Fig. 1). Cette différence perceptible même à l'oreille d'observateurs naïfs, l'est-elle par les marmottes? Pour tester cette hypothèse, 2 séquences artificielles de cris d'alarme ont été construites respec-tivement à partir de la triple répétition du cri d'un adulte ou d'un juvénile. La vitesse de répétition a été fixée à 0,6 s (variante la plus rapide observée chez cette mar-motte, reflétant la proximité d'un danger important) et la durée de chaque bloc à 4,5 s. Les niveaux d'intensité ont été équilibrés sur ceux d'un cri naturel avant le test. Les paramètres des sons émis (Fig. 1) sont les suivants :
| Cri adulte | Cri juvénile | |
| Durée du son, ms | 200 | 230 |
| Durée haute fréquence, ms | 80 | 100 |
| Étendue fréquence, kHz | 1.1-3.9 | 2.5-5.2 |
| Fréquence dominante, kHz | 3.6 | 4.5 |
| Amplitude sonore / 1 m du haut-parleur / dB | 95 | 95 |
Les enregistrements ont été diffusés (magnétophone à cassette Karpaty 205-1) lors des périodes d'activité maximum des marmottes (800-1200 h et 1600-2100 h). Celles-ci ont été observées, à la jumelle, de l'abri des bandes boisées bordant les pâturages et les cultures. La distance entre l'enregistrement de la réponse et la source sonore est inférieure à 100 m, et par vent violent, à 70 m. La distance maximum a été fixée par évaluation sub-jective de la bonne perception des signaux. La réponse d'un animal n'a été examinée qu'une fois par jour et au plus trois fois au cours de la période d'étude pour éviter l'habituation. Les marmottes n'étant pas marquées, pour ne pas tester le même animal soit on change d'animal visible soit de famille entre chaque essai. Le stimulus n'a été présenté que 30 mn ou plus après la première repasse, les différents blocs étant présentés de façon aléatoire. Le comportement des marmottes a été enre-gistré 4 minutes avant et après la repasse (seuls les ani-maux non alertés ont été pris en compte). La durée sépa-rant le stimulus de la réponse d'alarme (latence) et la durée de la vigilance, en distinguant lors des premières repasses les périodes suivantes : 0-30 s, 31-60 s, 61-90 s, 90-120 s et >120 s ont été chronométrées. De plus, l'âge, le nombre, la distance entre les animaux broutant ensemble, la distance par rapport aux terriers ont été notés. Après l'essai, la distance à laquelle se trouvait l'animal observé a été mesurée.
Cinq types de réponses, correspondant au niveau d'alarme, ont été définis d'après postures et mouvements : 1. absence de réponse, 2. bref regard vers la source du signal, 3. postures d'alarme spécifiques quadrupède ou bipède, 4. fuite vers le terrier, 5. fuite à l'intérieur du terrier. Les données ont été comparées à l'aide de statistiques non paramétriques, tests du Chi carré et de Fischer (programme Statgraf).
La réponse d'alar-me des jeunes en présence d'un ennemi potentiel et celle des adultes sont significativement différentes. Dans une famille, les jeunes présentent une activité vocale supé-rieure à celle des adultes et émettent plus souvent un cri d'alarme à l'approche de tout objet. Les premiers cris sont le fait d'un adulte, mais sont généralement terminés par les jeunes. Jamais 2 marmottes de la même famille ne crient simultanément en cas de faible danger : géné-ralement, le second émetteur ne crie que lorsque le pre-mier a cessé d'émettre, ou le premier cesse de crier lors-que le second animal commence à crier. Mais souvent 2 marmottes de différentes familles crient ensemble.
Les réactions de bobac face à différentes espèces de mammifères (chien, renard, lièvre, chevreuil, vache), d'oiseau de proie (vautour moine) et à l'homme ont été observées et enregistrées au magnétophone. Seuls les jeunes les considèrent toutes comme dangereuses : ils crient en présence de lièvres et même de vaches, bien que celles-ci soit l'élément le plus fréquent du milieu. Certains adultes sont, parfois, alertés par la présence d'un lièvre, mais ne crient jamais. Ils n'émettent pas de cris d'alarme face à un busard bleu, oiseau de proie moins imposant qu'un vautour, mais quand il se pose sur un monticule près des marmottes, les jeunes rentrent dans les terriers en silence et les adultes adoptent parfois la posture d'alerte quadrupède près de l'entrée du terrier. Mais toutes les marmottes quel que soit leur âge sont effrayées par le vautour, et émettent en sa présence un à trois cris d'alarme puis s'enfuient dans les terriers.
Lors d'approches lentes d'un observateur, la dis-tance moyenne de dérangement est la même pour les 2 classes d'âges : les marmottes commencent à crier entre 150-40 m de l'ennemi (moyenne 82 m). La distance de fuite vers les terriers varie : 6-8 m chez les jeunes et jus-qu'à 15-20 m pour les adultes. De plus, la probabilité de ressortir à la surface est significativement plus élevée chez les jeunes, que chez les adultes qui peuvent rester dans le terrier pendant des heures. Ces différences liées à l'âge suivent des variations saisonnières (sujet d'une prochaine publication). En préliminaire, plus les juvé-niles sont jeunes, plus leur activité vocale est élevée. La haute fréquence d'émission du cri d'alarme chez ces derniers est apparemment une caractéristique adapta-tive, facilitant la survie des marmottons inexpérimentés par le maintien d'un haut niveau de vigilance, même superflu, mais cette tendance entre en conflit avec le besoin d'une alimentation intensive et l'accumulation de la graisse nécessaire à l'hibernation.
II. Réponses au cri d'alarme.
Ce signal spécifi-que (ensembles de 30 s) a été présenté 95 fois à des adultes et 45 fois à des juvéniles et les différences entre leurs réponses sont nettes. Chez les adultes, la posture d'alerte quadrupède prévaut, alors que c'est la posture bipède chez les jeunes. Cette différence n'est pas impor-tante, car elle dépend non pas du niveau d'alerte mais de la taille de l'animal, les jeunes avec la bipèdie accrois-sent leur champ de vision. Aussi ces 2 postures ont été regroupées en un seul type (3). Dans l'ensemble, les jeunes s'alarment facilement, avec une forte probabilité de fuite, non significative, vers et dans un terrier (31% au lieu de 23% pour les adultes ; F = 1,01 ; df1 = 1 ; df2 = 138 ; P>0,05). Ils n'ignorent jamais le signal, alors que les animaux âgés n'alertent pas dans 6% des cas à la suite des repasses (Fig. 2). Et les jeunes restent en état d'alerte moins longtemps que les adultes ([[chi]]2=6,44 ; df=2 ; P< 0,05 ; Fig. 3) qui peuvent rester en posture d'alerte 30-40 mn à l'entrée du terrier alors que les juvéniles reprennent rapidement leur activité interrom-pue. Les latences des 2 classes d'âges sont voisines ([[chi]]2 =1,27 ; df=2 ; p> 0,07). Généralement, l'alarme est rapide, 80% des juvéniles et 75% des adultes répondent en moins de 2 s. La faiblesse de la durée de la vigilance des marmottons confirme le conflit, cité plus haut, entre maintien de la vigilance et alimentation. L'augmentation de l'activité sonore les premières semaines d'émergence, comme celle mise en évidence chez les jeunes écureuils terrestres, peuvent s'expliquer dans le cadre de la sélec-tion de parentèle, car l'avertissement de danger accroît avant tout la sécurité des proches, frères, soeurs et mère (Dunford 1977, Sherman 1985, Schwagmeyer 1980). Nous devrions sinon considérer que les propriétés du système nerveux des jeunes et leur absence d'expé-rience, conduisent à l'émission du cri d'alarme face aux objets sans danger.
III. Réponses aux variantes du cri d'alarme liées à l'âge.
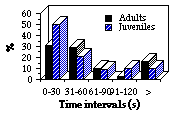
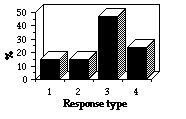
Soixante-douze repasses de cri d'alarme d'adultes et 68 de juvéniles ont été présentées 74 fois à des adultes et 66 fois à des jeunes pour tester leur capacité à distinguer le cri d'alarme émis par un adulte ou par un jeune. Aucun critère additionnel autre que sonore n'a été présenté dans les expériences. Aucune fuite vers le terrier n'a été observée, car contrairement aux repasses précédentes, la durée de présentation du stimuli est inférieure (4,5 s au lieu de 30 s), ce stimulus dérange moins longtemps les marmottes. Toutes les marmottes répondent de façon similaire, la seule diffé-rence marquante est que les animaux répondent aux signaux adultes presque deux fois plus souvent qu'à ceux des juvéniles, ils ignorent 8% des signaux adultes et 15% de ceux des juvéniles (Fig. 4-5). C'est la même chose pour chaque classe d'âge sans que les différences soient significatives ([[chi]]2 = 1,48 ; df =2 ; P> 0,3 pour les adultes ; [[chi]]2 = 0,19 ; df=2 ; P> 0,9 pour les jeunes). Les différences de durée de la réponse sont plus nettes. Ainsi, la moyenne de réponse des deux classes d'âge au signal d'adulte est de 55,9 s et celle au signal de jeune de 26,6 s (significatif au seuil 0,01 ; Fig. 6). Les réponses des adultes au signal adulte durent significativement moins que celles au signal des jeunes (dans le premier cas 70% des récepteurs restent alarmés plus de 20 s, et seulement 26%, plus de 80 s ; dans le deuxième cas, seulement 61% des marmottes sont en alerte plus de 20 s et 7% d'entre elles plus de 80 s). Chez les jeunes, 7% d'entre eux restent en alerte plus de 80 s suite à la variante adulte et aucun d'eux pour de telles durées en réponse à un signal de juvénile. Dans les 2 classes d'âge, les marmottes sont en alerte plus de 240 s en réponse au signal d'adulte seulement.
Comme les jeunes marmottes émettent générale-ment le signal d'alarme face à un danger potentiel peu différencié, contrairement aux adultes, et que les carac-téristiques spectrales de leurs cris diffèrent de celles des adultes, nous pouvons supposer que la valeur informa-tive plus faible de leur avertissement influence la réponse des récepteurs. Les tests confirment l'existence d'une discrimination des variations des cris d'alarme en fonction de l'âge chez M. bobac, comme cela a été montré dans les mêmes conditions chez la marmotte à longue queue (M. caudata). Chez cette dernière, la méfiance vis-à-vis des jeunes existe et la réponse au cri d'alarme dépend aussi de l'âge de l'émetteur (Nikol'skii & Nesterova 1990). Celle-ci est plus nette, probable-ment du fait de différences plus marquées du cri d'alar-me en fonction de l'âge. Aussi, comme l'avertissement d'une jeune marmotte est rarement fiable et l'alarme douteuse, la tendance de toutes les marmottes à s'alerter suite à des signaux de courte durée émis par les animaux âgés et à ignorer le plus souvent ceux émis par les jeunes, semble être justifiée biologiquement et béné-fique. Ainsi, elles prennent moins souvent en compte les nombreux cris d'alarme douteux au profit d'activités plus bénéfiques, comme l'alimentation. Les expériences démontrent aussi, la capacité des marmottes bobacs à reconnaître les paramètres spectraux des cris d'alarme. Il est intéressant de noter que nos recherches précédentes ont montré que les marmottes peuvent percevoir des différences plus fines de la structure spectrale du cri d'alarme spécifique en liaison avec la variabilité des signaux de situation, mais échouent à distinguer leur propre cri d'alarme de celui d'une espèce proche, M. baibacina (Nikol'skii & Nesterova 1988).
*****
| Figure 1. Sonagramme du cri d'alarme de M. bobac. Sonogram of M. bobac alarm call. | Figure 2. Réponses des marmottes au signal d'alarme.
Marmot responses to alarm signal.
|
 | 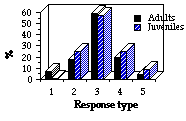 |
| Figure 3. Durée des réponses des marmottes au signal d'alarme. / Duration of marmot responses to alarm signal. | Figure 4. Réponses des marmottes au signal émis par un adulte. /
Marmot responses to adults' signal variant.
|
 |  |
| Figure 5. Réponses des marmottes aux signaux émis par les
juvéniles. / Marmot responses to juveniles' signal variant.
| Figure 6. Durée des réponses des marmottes aux signaux
émis par les adultes et par les juvéniles. / Duration of
marmot responses to adults' and juveniles' signal variants.
|
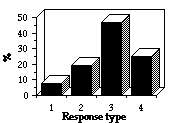 |  |
This report is part of a long-term investigation of marmots' ability to recognize their own alarm call between calls of related and distant (on the systematical level) species as well as to discriminate situation and age variability of conspecific alarm call, based on play-back field experiments. Three species, Marmota bobac, M. baibacina and M. caudata were studied (Nikol'skii, Nesterova, 1988, 1989, 1990, 1991). The aim of the study was to reveal age-dependant differences in mar-mot responses to conspecific alarm call, as well as to examine their sensitivity to the age cue of alarm signal spectral structure using field playback paradigms.
Responses of marmots to constructed acoustical stimulus were observed. With the help of a portable tape-recorder I broadcasted pre-recorded alarm calls, previously obtained in this very population, and artifi-cially arranged : 1) 30 s sets of sounds from natural alarm signal with 4 s, 2 s and 0.6 s variants of interval of sound repetition (constructed through repetition of a sound from alarm signal of an adult) to test differences between alert behaviour of adults and juveniles; 2) age variants of alarm signal to reveal whether age cue is essential to marmots. Alarm signal of M. bobac repre-sents a sequence of separate sounds (calls) with repeti-tion intervals from hundreds of ms to several s, and every sound consists of two components: a low and a high-frequency. Sonogram analysis showed that the latter has higher dominant frequency and longer dura-tion in juveniles' signal as compared with adults' one (Fig. 1). In the field even an inexperienced observer can recognize by ear whether a young marmot or an adult one is calling. Can marmots too? To examine this hypo-thesis, I constructed two artificial sequences of alarm calls, each consisting of 3 times repeated one sound of adult's or juvenile's signal, respectively. Repetition rate (sequence period) was 0.6 s (the most rapid variant of bobacs' signal, reflecting great and nearby danger). Duration of each block was 4,5 s. Intensity levels were calibrated equal to natural call prior to testing. Para-meters of transmitted sounds (Fig. 1) are as following:
| Adult's call | Young's call | |
| Sound duration, ms | 200 | 230 |
| High-frequency component duration, ms | 80 | 100 |
| Frequency range, kHz | 1.1-3.9 | 2.5-5.2 |
| Dominant frequency, kHz | 3.6 | 4.5 |
| Sound amplitude / 1 m from the speaker/ dB | 95 | 95 |
I broadcasted recordings using a cassette-tape recorder (Karpaty 205-1) during periods of maximum aboveground activity of marmots: (0800 to 1200 hours and from 1600 to 2100 h). Marmots were observed only from a shelter, forest stripes along pastures and agriculture fields. Distance of response registration was under 100 m from the signal source, and under strong wind conditions the distance was restricted to 70 m. Maximum distance was determined by my subjective assessment of good perception of transmitting signals.
Response of one animal was examined only once a day and three times at the most during the whole work period to avoid habituation. Marmots were not indivi-dually marked, but care was taken not to test the same animal by changing for a focal marmot or simply shif-ting to another family territory after each trial. The next stimulus was presented to the same marmot after 30 min. and more following the former stimulus play-back. I randomized the order of presentation of the different blocks. Marmots behaviour (only calm, not alerted animals were counted) was registered 4 minutes before and after play-backs. Duration between onset of stimulus presentation and onset of marmot alert reaction (latency of response) and duration of alertness, distinguishing the following time segments: 0-30 s; 31-60 s; 61-90 s; 90-120 s ; 120 s., were timed. Age, number of animals grazing together and their spacing, distance from the burrows were noted down as well. Observations were made with the help of binoculars. After the trial I measured distance from the observed marmot location.
Five types of responses corresponding to the level of alertness were defined by their alert postures and movements: 1 - no response; 2 - brief look towards the signal source; 3 - specific quadrupedal and bipedal alert postures; 4 - run to the burrow; 5 - run down into the burrow. We compared data using Chi-square and Fisher non-parametric statistics (Statgraf' program).
Significant differences of alarm behaviour in presence of potential enemy was obtained between adults and juveniles. When family en-counted a dangerous object, juveniles showed higher vocal activity, than nearby adults and emitted alarm call with higher probability to any approaching object. Although the first calls may belong to an adult family member, alarming bout was usually finished by a juvenile. Two marmots from the same family never called simultaneously when danger is moderate: usually the second alarmer began calling only when the former one stopped or otherwise the former one used to stop calling if the second animal started. But two marmots from different family called together quite commonly.
Alarm reaction of bobacs to several species of mammals: dog, fox, hare, roe, cow; to bird of prey: black kite; and to man have been observed and recor-ded. Only juveniles considered all of them as dangerous. They called to hares and even to cows, although grazing cattle was the most common component of their landscape. Some adults were alerted to a certain extent by a running hare, but they did not call. I did not obser-ve marmots calling to a hen harrier - less than a kite but large enough prey-bird, but if it landed on a mound close to marmots, juveniles silently enter a burrow, and adults sometimes alerted near burrow entrance in a quadrupedal alert posture. But all marmots indepen-dently on age were frightened of a black kite, and at once gave 1-3 alarm calls and escaped into burrows.
Slow approaches of marmots by an observer showed that the disturbance distance was the same for both ages, calling started at 150-40 m from the enemy, mean value 82 m. But the distance, when marmots ran away into a burrow, varied. Juveniles allowed observer to approach them up to 6-8 m., and adults up to 15-20 m before escaping. Moreover, probability of re-emerging on the surface was significantly higher in juveniles, while disturbed adults could stay down the burrow even for hours. It is important to mention that age differences in marmots alert behaviour undergo seasonal variations, but it is a subject of a next paper. Preliminary it is to say, that the younger the juveniles are, the higher their vocal activity is. High frequency of emitting alarm call in juvenile marmots is apparently an adaptive feature, which helps inexperienced cubs to survive through maintaining high vigilance level, even superfluous, but this tendency conflicts the need of intensive feeding to accumulate necessary amount of fat for hibernation.
II. Responses to alarm call. Conspecific alarm signal (30 s duration sets) was presented to marmots 140 times (95 to adults and 45 to juveniles). The results revealed significant differences in responses of adults and juveniles to the alarm call . In adults quadrupedal alert posture prevailed, on the contrary juveniles more often alerted bipedal. But I think that these differences are not important, because they depend not on alert level, but only on animal size, smaller young need to stay bipedal to enhance their range of vision. So data on bipedal and quadruped postures were pooled in the same posture type. Juveniles on the whole tended to alert strongly, showed higher but not significant proba-bility of run to and down into a burrow (31% versus 23% in adults; F = 1.01; df1 = 1; df2 = 138, P>0,05). However, juveniles never ignored the signal, whereas elder animals did not alerted in 6% of play-backs (Fig. 2). But juveniles stayed alerting for less period, than adults ([[chi]]2=6.44; df=2; P< 0.05; Fig. 3). In some cases adult marmots stayed in alert posture at burrow entrance for 30-40 minutes following a stimulus presen-tation. Juveniles never delayed at the mounds and quicker resumed interrupted activity. Latencies of both age groups appeared similar ([[chi]]2 =1.27; df =2; p> 0.07). Almost in every trial marmots were alerted immediately, 80% of juveniles and 75% of adults responded under 2 s. Less duration of cubs alertness proved above mentioned conflict of tendencies to keep permanent vigilance versus spend more time for feeding activity. Explanations of increased acoustic activity in the first weeks after emerging, as was shown for example for the juvenile ground squirrels may be found in frames of the "kin selection" conception, since the warning about danger rises safety first of all of the close relatives, sibs and mother (Dunford 1977, Sherman 1985, Schwagmeyer 1980). Otherwise we should consider the peculiarities of the cubs' nervous system and lacking life experience, that leads to calling to objects which are not dangerous at all.
III. Responses to age-variants of alarm call.
Seventy two play-backs of adult's alarm signal variant and 68 of juvenile's were presented to adult marmots 74 times and 66 times to young in experiments concerning marmots' ability to recognize whether an adult animal is calling or a young one. No additional cues, except acoustical were presented in the experi-ments. In all the trials no escapes into the burrows were observed, because unlike previous play-back experiment in this one the duration of presented stimuli was much less (4,5 s, compared to 30 s), so the stimuli disturbed marmots less . Thus I compared 4 posture types. It was discovered that all marmots responded quite similarly, the only striking difference is that animals did respond to adult's signal nearly twice more often than to the juvenile's, i.e. marmots ignored 8% of adult's signal and 15% - of juvenile's (Fig. 4-5). This is similar in both age groups of recipients. But differences are not significant ([[chi]]2 = 1.48 df =2 P> 0.3 for adults; [[chi]]2 = 0.19 df=2 P> 0.9 for young). Differences in response duration appeared much more significant. Thus, joint mean response of both age groups to the adult's variant amounts to 55,9 s and to juvenile's - 26,6 s (differences are significant at the level p< 0.01) (Fig. 6). If consider the results separately: responses of adults to adult's variant were significantly of less duration, than to the juvenile's one - in the former trials 70% of recipients stayed alerted over 20 s, and 26% - even over 80 s, and in the latter trials only 61% of marmots were alerted more than 20 s and 7% - more than 80 s; as for young, 7% of them stayed alerted to adult's variant over 80 s, and no one for such a long period in response to juvenile's signal. In both age groups marmots were alerted up to 240 s (and more) only in response to adult's variant.
Since young marmots usually give alarm signal in presence of potential danger less differentiated, than adults, and even to threatless objects, and that spectral features of their calls differ from those of adults we could suppose, that their warning of lowest informa-tional value influences the response behaviour of reci-pients. Conducted tests confirm the supposition that bobac marmots can discriminate their alarm signal age-variants. I would like to mention, that these results are in agreement with those obtained in similar field experi-ments with long-tailed marmots: distrust of the young took place in that case too and long-tailed marmots responded to alarm call as well in dependence on the alarmer's age (Nikol'skii & Nesterova 1990), and this expressed even clearly probably because of greater difference between the species alarm call age variants. So, because the warning of a young marmot often prove to be incorrect and that the alarm may appear unre-liable the tendency of marmots, both adults and young, to alert for significantly shorter period to signal from elder animals and to ignore comparatively more often the one from the young seems to be biologically justified and beneficial for marmots. By this way they may pay less attention to numerous unsuitable alarm calls in favour of other forms of positive activity, for example, foraging. Experiments also demonstrate high enough ability of bobac marmots to recognize spectral parameters of such acoustical signal of their vocal repertoire as alarm call. It is interesting to mention, that our earlier researches have shown, that marmots can perceive much more slight differences in conspecific alarm call spectral structure, connected with signal situational variability, but fail to discriminate their own alarm call versus the one of closely related species - M. baibacina (Nikol'skii & Nesterova 1988).
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |