![]()
Le cri d'alarme des marmottes eurasiennes a été enregistré
sur le terrain. La dynamique spectrale (sonagramme) est spécifique alors
que l'organisation rythmique ne l'est pas. Cette dernière, correspondant
au relief, présente des parallélismes interspécifiques et
une divergence intraspécifique. L'augmentation de la division verticale
du relief s'accompagne d'une réduction des périodes des
répétitions d'impulsions. L'analyse spectrale suggère que
M. caudata et M. menzbieri ont été isolées
précocement des autres espèces. La dynamique spectrale du cri
d'alarme de M. camtschatica a des caractéristiques
intermédiaires entre celles des espèces eurasiennes et
américaines. La variabilité interspécifique de la
structure du cri d'alarme chez M. sibirica, M. baibacina et M.
bobac présente les caractéristiques d'un cline graduel. Les
signaux de ces trois espèces (groupe "bobac") ont une composante de
basse fréquence similaire. La structure du signal de M. marmota
ressemble plus à celle du signal des marmottes américaines.
Mots-clés : Marmotte grise, Marmota baibacina, Marmotte des steppes, Marmota bobac, Marmotte à tête noire, Marmota camtschatica, Marmotte rouge, Marmota caudata, Marmotte de Menzbier, Marmota menzbieri, Marmotte de Mongolie, Marmota sibirica, Eurasie, cri d'alarme, caractéristiques spécifiques, parallélisme interspécifique.
The alarm call of Eurasian Marmots was tape-recorded in field condition. The
dynamic spectrum (sonogram) is species specific whereas the rhythmic
organization is not. The rhythmic organization corresponding to the landscape
relief manifests the interspecies parallelisms and intraspecies divergence. The
increase of the vertical division of the relief is accompanied by reduction of
the periods of pulse recurrence. The analysis of dynamic spectra suggests that
M. caudata and M. menzbieri were isolated earlier than other
species. The alarm call dynamic spectrum in M. camtschatica has
intermediate features between those of Eurasian and American species. The
interspecific variability of the structure of alarm call in M. sibirica,
M. baibacina and M. bobac bears the character of stepped cline.
The signal of these three species ("group bobac") has similar low-frequency
component. M. marmota signal is more similar by its structure to the
signal of american species .
Key-words: Grey marmot, steppe marmot, black-capped marmot, red marmot, Mongolian marmot, Eurasia, alarm call, species specificity,
interspecies parallelisms.

Ce travail est une compilation des travaux de l'auteur (Nikol'skii 1969, 1974, 1976, 1984, Nikol'skii & Orlenev 1980, Nikol'skii et al. 1983). Les dernières données obtenues, soulèvent de nouvelles questions, mais ne remettent pas en cause et corroborent même les principales conclusions des travaux antérieurs.
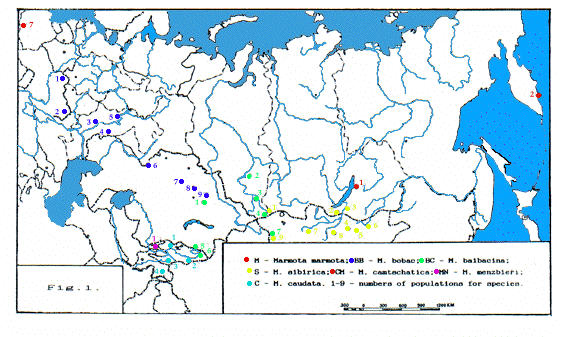
Le cri d'alarme (signal) est un caractère biolo-gique typique des Marmotinae : marmottes, écureuils terrestres et chiens de prairie. L'analyse du signal mon-tre que de nombreux paramètres sont spécifiques et pourraient être utilisées comme critères de reconnais-sance des espèces. Des analyses inter et intraspécifiques du signal de 7 espèces ont été réalisées, soit l'ensemble des espèces eurasiennes à l'exclusion de M. himalayana. Les signaux ont été enregistrés au magnétophone, uni-quement sur le terrain. Les données ont été collectées pendant 30 ans par de nombreux zoologistes I. Baklushingkaya (Yanina), N. Formozov, L. Kotsiyan (Slovaquie), T. Lisitsina, N. Nesterova, D. Orlenev, O. Pereladova (Novikova), M. Rutovskaya, M. Sukhanova, A. Nikol'skii. Les marmottes bobac (M. bobac), grises (M. baibacina), de Mongolie (M. sibirica) et à longue-queue (M. caudata) sont représentées par de nom-breuses populations. Les enregistrements sont conservés à la Collection des Sons de Mammifères du département de biologie de l'Université de Moscou. Les méthodes, les régions, les modes de collection des données et leur volume ainsi que les résultats ont été publiés antérieure-ment (bibliographie). Pour donner une idée de l'étendue géographique de la recherche, seules les principales régions de collection sont citées (Fig. 1) : 1) M. marmota : Tatras ; 2) M. bobac : Ukraine, région de la Volga, Kazakhstan ; 3) M. baibacina : montagnes du Kent (Kazakhstan central), Tien Shan, Sibérie ouest, Altai, Altai de Mongolie ; 4) M. sibirica : Tuva, région du Transbaikal, Mongolie ; 5) M. camtschatica : environs du lac Baikal, Kamtchatka ; 6) M. caudata : Tien Shan, Pamirs ; 7) M. menzbieri : Tien Shan.
Le signal de marmotte est formé de sons distincts groupés en séquences temporelles, ensembles ou séries. Chaque son a un spectre et une modulation de fréquence typiques. Le signal est représenté par deux groupes de caractéristiques; les structures rythmique et spectrale. Leur analyse montre que la structure spectrale est spéci-fique, alors que l'organisation rythmique est parallèle dans différentes espèces et sujette à une variabilité géo-graphique sous l'action de différents facteurs du milieu. L'organisation rythmique du signal est en corrélation avec la visibilité du paysage, mesurée par le morcelle-ment du relief. Cette liaison entraîne des parallélismes interspécifiques et des divergences intraspécifiques de l'organisation rythmique. Les principaux types de relief des habitats de marmottes, des plaines et collines faible-ment escarpées aux reliefs montagneux fortement morcelés, résultent du ravinement des versants et du plissement de leur surface, ce qui masque à l'obser-vateur la perspective lointaine (Figure 2). A chaque type de relief correspond une organisation rythmique parti-culière du signal. Dans les cas extrêmes, nous observons des ensembles de sons lents (plaines ou collines) et des séries de sons rapides (hautes montagnes découpées). Dans les cas intermédiaires, soit des ensembles de sons double ou triple (fig. 2), soit des séries de sons lents (non figurés) peuvent apparaître. Chaque type de struc-ture rythmique correspond à une valeur modale de la période de répétition sonore (période séquentielle). Ce paramètre est en corrélation avec le morcellement vertical du relief (fig. 2).
* Le signal de M. bobac, habitant les plaines et les collines basses, est formé uniquement d'ensembles de sons à succession lente. Ce type de signal apparaît aussi chez certaines populations (M. baibacina) vivant dans les zones au relief faiblement escarpé, et très rarement chez les populations de M. sibirica.
* Chez M. sibirica, des plateaux du Transbaïkal et de Mongolie, des ensembles de sons double ou triple, sont observés, comme quelquefois chez M. baibacina.
* Les séries de sons à succession rapide sont typiques de M. caudata et de la marmotte de Menzbier, et apparaissent dans les populations de marmottes grises des zones de haute altitude au relief fortement morcelé.
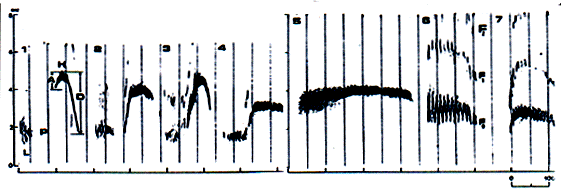
Ainsi, nous voyons que les parallélismes parti-culiers de l'organisation rythmique du signal relèvent des parallélismes écologiques et ne sont pas spécifiques. Au contraire, la structure spectrale du signal est spécifi-que et constitue un critère sûr de détermination. Les sonagrammes du cri d'alarme des espèces étudiées (Fig. 3) montrent que chaque espèce est aisément recon-naissable, quelle que soit l'origine de la population, par de nombreux critères qualitatifs diagnostiques et, on doit le souligner, par ces seuls critères. En effet, les paramètres quantitatifs des différentes espèces, telle que fréquence et durée des sons, se chevauchent largement
Nous allons décrire les critères qualitatifs spécifi-ques, et les quantitatifs qui peuvent varier avec des modifications situationnelles et d'âges, mais aussi avec la variabilité géographique, de chacune des espèces.
* Le signal de M. bobac. est formé d'une séquence de sons, dont chacun comprend deux composantes spec-trales, l'une à basse fréquence (L) et l'autre à haute fré-quence (H), séparées par une pause (P, fig. 3). Les fré-quences ascendante et descendante (branches des carac-téristiques de la modulation de fréquence, c'est-à-dire, les sonagrammes) sont asymétriques : la fréquence ascendante (A) est moins étendue que la fréquence descendante (D, Fig. 3). Le coefficient d'asymétrie (IA =A/D) n'excède pas 1 pour l'espèce sur toute son aire de distribution (Nikol'skii et al. 1983).
* Chez M. baibacina, comme chez M. bobac, deux composantes spectrales sont typiques. Mais contraire-ment à M. bobac, la composante à haute fréquence présente un canevas d'asymétrie inverse : la fréquence ascendante est plus étendue que la descendante. Et le coefficient d'asymétrie (IA =A/D) est supérieur à 1 pour toutes les populations étudiées.
* Chez M. sibirica, comme chez les précédentes, le signal présente deux composantes spectrales, mais la pause entre les deux est absente. La basse fréquence s'arrête juste avant que la haute fréquence commence ou, dans une certaine mesure, les deux composantes se chevauchent.
* Chez M. camtschatica, la composante à basse fréquence est présente mais très faible, concentrée dans une étroite bande de fréquences. Il n'y a pas de pause entre les deux composantes.
* Chez M. menzbieri, les sons ont une durée comparati-vement plus importante. La modulation de fréquence est faible. Presque toujours, le début du cri est précédé par un élément, qui est homologue, selon nous, à la composante de basse fréquence.
* Chez M. caudata, le signal est le plus distinct. Chaque cri représente un trille, s'élevant en résultat de l'amplitude de modulation.
* Chez M. marmota, la caractéristique la plus typique est l'absence absolue de la composante de basse fréquence.
Dans les signaux de toutes les espèces de mar-mottes, des harmoniques plus ou moins prononcées sont présentes (F1, F2, fig. 3). La fréquence dominante, portant l'énergie maximum, est concentrée dans la fréquence fondamentale (F0). L'analyse de la variabilité géographique des signaux de M. sibirica et de M. camtschatica nécessite une étude complémentaire. Le rapport des fréquences ascendante et descendante de la composante à haute fréquence du signal de M. sibirica montre une forte variabilité : d'une dominance de la fréquence ascendante sur la descendante jusqu'à leur égalité (canevas symétrique de la modulation de fréquence). Le rapport temporel des composantes de basses et de hautes fréquences varie aussi, d'un faible chevauchement à une fusion presque totale. De plus, le signal de cette espèce présente une grande variété d'harmoniques pour les deux composantes de fré-quences. Chez la marmotte à tête noire, l'étude de la variabilité géographique de la composante à basse fréquence et la symétrie/asymétrie des fréquences ascendantes et descendantes reste à faire. Selon les tra-vaux antérieurs (Nikol'skii et al. 1991), nous avons l'impression que les populations de l'Est de cette espèce diffèrent de celles de l'Ouest par l'absence de la compo-sante à basse fréquence. Mais, le sonagramme du signal d'une marmotte du Kamtchatka (Cap Shipunskii, l'un des sites les plus orientaux de l'aire de distribution de cette espèce), corrobore l'existence de la composante à basse fréquence dans les populations orientales aussi.
L'analyse des sonagrammes permet d'émettre les conclusions et hypothèses suivantes sur les inter-relations entre marmottes en ce qui concerne la systématique et leur aire de distribution.
1. Les signaux de M. bobac, M. baibacina, M. sibirica, du groupe "bobac", ont une caractéristique commune, une composante à basse fréquence, ce qui confirme leur parenté génétique. Cependant, le signal de bobac diffère de celui de la marmotte grise par l'asymétrie de sa composante à haute fréquence, et celui de la marmotte tarbagan diffère de celui des autres espèces par l'absence de pause entre les deux composantes.
2. Le signal de la marmotte à tête noire diffère par une faible composante à basse fréquence, un canevas symé-trique ou asymétrique de la modulation de la compo-sante à haute fréquence et par sa durée relativement plus longue. Ces caractéristiques sont intermédiaires entre celles des espèces du groupe bobac et celles des espèces américaines.
3. Le signal de M. menzbieri et plus particulièrement celui de M. caudata se distinguent de ceux des espèces précédentes, supportant l'hypothèse d'un isolement ancien des aires de distribution de ces espèces. Mais la présence d'une composante à basse fréquence rapproche M. menzbieri des espèces du groupe "bobac".
4. La structure du signal de M. marmota est voisine de celle des espèces américaines, en particulier par l'absen-ce de la composante à basse fréquence (le son ressem-blant à un sifflet). Cette caractéristique rapproche, formellement, M. marmota des espèces américaines.
5. La variabilité interspécifique de la structure du signal au sein du groupe "bobac" suit un cline graduel, caracté-risé par un raccourcissement de la pause entre compo-santes à basse et haute fréquence de l'Est à l'Ouest. De plus, l'asymétrie de la composante à haute fréquence du signal de M. sibirica correspond à celle du signal de M. baibacina (espèce géographiquement proche), et non à celle de l'espèce distante, M. bobac. Ces faits fournis-sent militent en faveur de la formation d'unités géo-graphiques par suite de l'alternance de périodes de panmixie et d'isolement géographique.
6. L'analyse de la variabilité spatiale du signal a aussi révélé que l'isolement géographique affecte la variabi-lité intraspécifique de M. caudata. La durée des sons de la séquence sonore diffère entre les populations du Tien Shan et d'Alai et celle des populations de l'Est du Pamir. Dans cette dernière zone, les sons deviennent significa-tivement plus courts du début de la série à la fin. L'Est Pamir, aujourd'hui comme au Pléistocène, représente en effet une zone interglaciaire isolée par les glaciers. L'isolement des populations l'occupant est probablement la raison de la variabilité géographique du signal (Nikol'skii & Orlenev 1980). Cette variabilité chez la marmotte à longue-queue a été observée pour la pre-mière fois par Bibikov, il y a une quarantaine d'années.
Ainsi, l'analyse de la variabilité du signal d'alar-me de la marmotte indique que ses caractéristiques sont influencées par : 1) des corrélations génétiques entre les espèces ; 2) l'histoire et la structure des aires de distribu-tion ; 3) des facteurs du milieu. Différents critères du signal sont contrôlés par différents facteurs, ce qui conduit à une variabilité en mosaïque : spécifique pour certains critères et parallélisme interspécifique pour d'autres. Ces derniers confirment la règle des séquences homologues dans la variabilité héréditaire de Vasilov et sont la confirmation indirecte que les différents para-mètres d'un signal (structure spectrale et organisation rythmique) sont contrôlées par différents gènes.
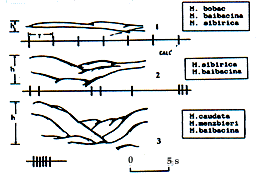
| Morcellement du relief et types d'organisation temporelle du cri d'alarme : 1.
zone plate ou collines faiblement escarpées / ensembles de cris
séparés ; 2. moyenne montagne peu décou-pée /
ensembles de cris doublés ou triplés ou séries lentes ; 3.
haute montagne fortement découpée / séries rapides. T :
période d'une séquence de cris ; h : ravinement du relief. Forms of the vertical dismemberment of the relief and corresponding types of the alarm call's temporal organisation: 1. flat, hilly slightly dismembered countries / sets of separate calls; 2. low-mountain middling dismembered countries / sets with doubled-tripled calls or slow series; 3. tall mountain deeply dismembred countries / rapid series. T: sequence periods of calls; h: relief's vertical dismemberment.
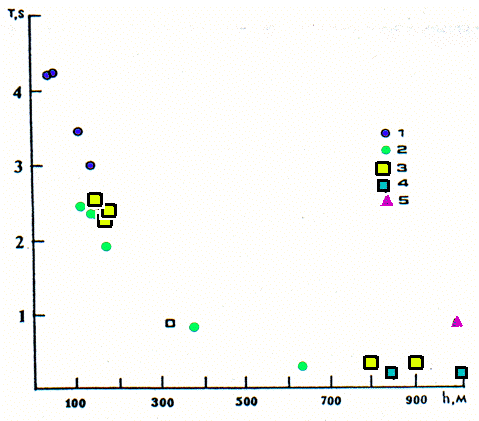
| Dépendance des périodes séquentielles des cris (T)
au morcellement du relief (h). Dependence of sequence periods of calls (T) on the vertical dismemberment of relief (h).
1. M. bobac ; 2. M. baibacina ; 3. M. sibirica ; 4. M. caudata ; 5. M.
menzbieri |
This paper represents a compilation of author's works published in Russian (Nikol'skii 1969, 1974, 1976, 1984, Nikol'skii & Orlenev 1980, Nikol'skii et al. 1983). New large data have been obtained during last years, that put several new questions, but did not influenced principally the conclusions set in earlier publications and corroborate the main of them.
Alarm call (or signal) is a typical form of biology of Marmotinae - marmots, ground squirrels and prairie dogs. The signal analysis shows that many of its para-meters are species-specific and could be used in species recognition. We have conducted inter and intraspecific analysis of the signal of 7 Eurasian marmot species which represent all the Eurasian ones, M. himalayana excepted. Signals were tape-recorded in field conditions only. Data have been collecting for 30 years by a large group of zoologists: I. Baklushingkaya (Yanina), N. Formozov, L. Kotsiyan (from Slovakia), T. Lisitsina, N. Nesterova, D. Orlenev, O. Pereladova (Novikova), M. Rutovskaya, M. Sukhanova, A. Nikol'skii. Bobac, grey, Mongolian and long-tailed marmots are represented by a large number of populations. Tape-recordings collection is kept in the Mammals Sound Collection at the Biology Department, Moscow University.
In order not to take up place I do not describe methods, regions of data collecting and data volume in details and do not list publications, except the works this paper is based on. All results are in details presen-ted in previous authors publications given in the refe-rence list. I would mention only the main geographical regions of data collecting, that gives a general idea on geographical range of conducted research: 1) Marmota marmota - Tatras; 2) M. bobac - Ukraine, Volga region, Kazakhstan; 3) M. baibacina - the Kent Mountains (Central Kazakhstan), Tien Shan, West Siberia, Altai, Mongolian Altai; 4) M. sibirica - Tuva, Transbaikal region, Mongolia; 5) M. camtschatica - the lake Baikal environs, Kamtschatka; 6) M. caudata - the Tien Shan, Pamirs; 7) M. menzbieri - Tien Shan (Fig. 1).
Marmot signal consists of separate sounds grouped in temporal sequences, sets or series. Every separate sound has got typical spectre and frequency modulation. On the whole, the signal is represented by two groups of characteristics - the rhythmical and spectral structures. Temporal and spectral analysis of the signal shows, that its spectre structure is species-specific, whereas the rhythmical organization is parallel in different species and subjected to geographical varia-bility under different environmental factors. Signal rhythmical organization correlates with the visibility of landscape, which relativity can be measured through the relief vertical dismemberment. Connection of the signal structure with an environment results in interspecies parallelisms and intraspecies divergence of its rhythmical organization.
Figure 2 shows main relief types of marmot habitats - from slightly dismembered plain or a hilly relief with small vertical dismemberment to deeply dismembered high-mountain relief. First of all the result of the incises in main slope and the folding surface, is that far perspective is hidden from an observer. To every type of relief corresponded a particular rhythmi-cal organization of the signal. In the outermost cases we observed sets of slowly emitted sounds (plain or hilly relief) and series of rapidly emitted sounds (high-mountain deeply dismembered relief). In the inter-mediate cases there may appear either sets of doubled-tripled sounds, as shown in the figure, or series of slowly emitted sounds, not shown in the figure.
Every type of rhythmical structure is in corres-pondance with one or another modal value of a period of sound repetition (sequence period). This parameter correlates with a relief vertical dismemberment, as it is demonstrated in figure 2:
- Signal of bobac marmot, inhabiting plains and low hills, consists of sets of slowly following sounds only. This signal also occurs in some populations of grey marmot (M. baibacina) inhabiting areas with slightly dismembered relief, and very seldom in that of tarbagan (M. sibirica).
- For tarbagan inhabiting plain plateaux of Transbaikal region and Mongolia, sets of doubled-tripled sounds sometimes also observed in grey marmot, are typical.
- Series of quickly following sounds are only typical for the mountain long-tailed marmot and Menzbier's marmot and occur in populations of grey marmot in high-mountain lands with deeply dismembered relief.
So, we see that parallelisms are peculiar to the signal rhythmical organization and this rouses from ecological parallelisms and can not be a species sign. On the contrary, signal spectral structure is species-specific and serves as a reliable determining sign.
Sonograms of alarm signal of the species (fig. 3) show that the signal of every species is easily distin-guished, irrespective of its population origin, by several diagnostic qualitative signs and, it should be empha-sized, only by qualitative ones. Because quantitative characteristics, such as frequency and duration of sounds, may broadly overlap in different species.
Below, signal of every of the 7 species is described : species specific qualitative signs and quanti-tative characteristics which may vary with situational and age changes, as well as geographical variability.
* M. bobac. Signal consists of a set of sounds, each includes two spectral components, low (L) and high-frequency (H) ones, divided by a pause (P, fig. 3). Ascending and descending frequencies (branches of the frequency modulation characteristics, i.e. sonograms) are asymmetrical: the ascending frequency takes up much less frequency range, than that the descending one (A and D, fig.3). Coefficient of asymmetry (IA = A/D) does not exceed 1 all over the species area (Nikol'skii et al. 1983).
* M. baibacina. Similar to the former species, two spectral components are typical for grey marmot signal too, but unlike that of bobac marmot high-frequency component shows inverse asymmetry pattern - the ascending frequency takes up much more frequency range, than the descending. And coefficient of asymmetry (IA =A/D) on the contrary is larger than 1 in all examined populations.
* M. sibirica. Similar to the former two species, two spectral components are also present in the signal of Mongolian marmot (tarbagan), but unlike the former two species there is no pause (P) between low-and high-frequency components. The low-frequency component ends just before the high-frequency one starts or both components may overlap to some extent.
* M. camtschatica. The low-frequency component is present, but it is very slight and concentrates in narrow frequency range. There is no pause between the low- and high-frequency components.
* M. menzbieri. Comparatively long duration of sounds. Slight frequency modulation. Almost always the call beginning is preceded by an element, that is homolo-gous in our opinion to a low-frequency component.
* M. caudata. Signal of this species is the most unique one. Every call represents a trill, arising as a result of amplitude modulation.
* M. marmota. The most typical feature of European marmot signal is absolute absence of low-frequency component.
In the signals of all marmot species more or less pronounced harmonics are present (F1, F2, fig. 3). Dominant frequency, carrying maximum of energy, is concentrated in the fundamental frequency (F0).
Analysis of geographical variability of tarbagan and black-capped marmots signals needs further exami-nation. Ratio of the ascending and descending frequen-cies of the high-frequency component in tarbagan signal shows great variety - from significant predominance of ascending frequency versus the descending one up to their equality (symmetrical pattern of frequency modu-lation). Temporal ratio of the high and low-frequency components varies too from poor overlapping to nearly absolute merging. Moreover, in this species signal a wide range of variants of harmonics expression for the both frequency components can be observed. In black-capped marmot investigation of geographical variabi-lity of the low-frequency component expression and symmetry/asymmetry of the ascending and descending frequencies is needed. According to the earlier publi-shed work (Nikol'skii et al. 1991), it may appear an impression that absence of a low-frequency component differs east populations of the species from the west. Figure 3 shows sonogram of this marmot signal from Kamtschatka (Shipunskii cape - one of the most far east sites of the species area), corroborating that the low-frequency component is present in east populations too.
Sonogram analysis permits to propose following conclusions and hypothesis on marmots interrelations regarding systematic and their areas forming:
1. Marmots of the "bobac group" - M. bobac, M. baibacina, M. sibirica - carry a common group feature, their signals include a low-frequency component, that confirms the opinion about their genetic relationship. However, bobac marmot signal differs from that of grey marmot by a high-frequency component asymmetry, and tarbagan marmot signal differs from the two former by lacking of an interval between the low-and high-frequency components .
2. Black-capped marmot signal differs by the faintness of low-frequency component, symmetry or asymmetry pattern of the high-frequency component frequency modulation and its comparatively long duration. Typical features of black-capped marmot signal, as we regard, take an intermediate place between those of the "bobac group" and American species, which typically have no low-frequency component.
3. Menzbier's marmot signal and particularly that of long-tailed marmot most of all differ from the former four species signals, that leads to supposition about their earlier isolation during forming of the species areas. But presence of a low-frequency component likens Menzbier's marmot to "the bobac group" species.
4. European marmot signal is more similar by its structure to the signal of American species, that finds its expression in lacking a low-frequency component (the pure whistle-like sound). Formally this feature likens the European marmot to American species.
5. Interspecies variability of the signal structure within the "bobac group" has got a pattern of graded cline, that finds expression in shortening of a time interval between the low- and high-frequency components from west to east direction. Moreover, an asymmetry of the high-frequency component in tarbagan signal corres-ponds to the asymmetry of signal of its area neighbour - grey marmot, but not to that of spatially distant bobac marmot. These facts set grounds for discussing of forming of units for the group area as an alternation of panmixia periods with geographical isolation.
6. Analysis of the signal spatial variability has also revealed that geographical isolation affected intra-species variability of long-tailed marmot. Tien Shan and Alay populations of this species differ from those in East Pamir by duration of sounds in the signal sequence: in the latter area sounds turn significantly shorter from the series beginning to end. We refer this to the fact, that East Pamirs today as well as in the Pleistocene has represented an interglacial zone being isolated by glaciers. Isolation of marmot population inhabiting the area is likely to be a reason of the signal geographical variability (Nikol'skii & Orlenev 1980). For the first time geographical variability of long-tailed marmot signal was noticed by professor D. Bibikov about 40 years ago. Thus, analysis of marmot alarm signal varia-bility indicate, that its characteristics are influenced by: l) genetic interrelations among species; 2) history and structure of areas; 3) environmental factors.
Different signal features are controlled by diffe-rent factors, that leads to mosaic of variability - species specificity of one features and interspecies parallelisms of others. The latter as a phenomenon agrees with the Rule of Homologous sequences in hereditary variability according to N.I. Vavilov and is an indirect confirmation of the fact, that different characteristics of the same signal (spectral structure and rhythmical organization) are controlled by different genes.
A suivre
Forward ![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |