![]()
Les recherches sur la biologie reproductive et l'hibernation des marmottes des
steppes (M. bobac) en captivité sont présentées
dans ce travail. L'analyse des pertes pondérales des marmottes
hibernantes amène à l'hypothèse que l'arrêt de
l'hibernation est étroitement associé aux dépenses
énergétiques de reproduction des animaux. Celle-ci débute
avec l'élévation de la température corporelle et dure 2
à 3 semaines. Puis la température atteint 36-38deg.C, la
reproduction proprement dite commence alors. Les retours à l'hibernation
au printemps ne se produisent que chez les animaux très faibles chez qui
la période pré-reproductive continue quelque temps. Mais leur
niveau reproducteur n'atteint jamais un seuil suffisant pour permettre leur
reproduction.
Mots-clés : Marmotte des steppes, Marmota bobac, hibernation, température
corporelle.
The results of many years of investigation on breeding biology and
hibernation of steppe marmots in captivity are presented in this work. Analysis
of weight losses of hibernating marmots lead to suppose that the end of
hibernation is closely connected with the expense of energetic resources of
breeding animals. The breeding process starts simultaneously with the increase
of body tem-perature and lasts 2 - 3 weeks. After that temperature of animals
increases to a constant (36-38deg. C) and breeding itself begins. Returns to
hibernation in spring after a period of starvation was recorded only in very
weak animals among which prebreeding period continues some time. But their
reproductive index never reaches a suitable level for successful
breeding.
Key-words: Steppe marmot, Marmota bobac, hibernation, body temperature, breeding.

La température corporelle des marmottes change lors de l'arrêt de croissance pondérale. En été, des températures élevées et constantes (36-38deg.C) sont enregistrées chez toutes les marmottes adaptées, puis elles commencent à fluctuer entre 27-38deg.C lorsque la masse maximum est atteinte (août). A cette période, un arrêt d'alimentation ou des températures ambiantes trop froides peuvent entraîner une chute de la température corporelle et le passage à l'hibernation.
La première période d'hibernation vraie en éle-vage débute en septembre après l'arrêt de l'alimentation. Le poids moyen des animaux reproducteurs est alors de 6,1 kg pour les femelles et de 7,2 kg pour les mâles. Aucun individu dont le poids a été inférieur à 4,9 kg à la mi-septembre n'a été reproducteur (Tab. 1). Les pertes de poids des marmottes reproductrices au cours de cette période (mi-septembre-début novembre) sont importantes (13,5% en moyenne de la masse totale lors des 40 premiers jours, soit 3,4 g par kg par jour). La tempéra-ture corporelle moyenne décroît de 32 à 14deg. C. Le poids minimum de ces marmottes, début novembre, a été de 4,4 kg (Tab. 1). Les pertes de poids durant la période d'hibernation "profonde" (novembre à mi-janvier) sont minimales (1,3 g/kg/j. L'indice minimum de masse corporelle des animaux reproducteurs est de 3,8 Kg pour les femelles et de 4,1 kg pour les mâles à la mi-janvier (Tab. 1). La dynamique de la température corpo-relle durant l'hibernation profonde varie suivant les individus ainsi que le nombre et les moments des réveils. Ces derniers ne dépendent pas de la température ambiante et la température moyenne des animaux est de 9,7deg.C. Plus une marmotte a une température moyenne élevée et des réveils fréquents, plus ses pertes de poids sont importantes. Par exemple, elles sont de 3,1 g/kg/j chez une marmotte non reproductrice dont la température moyenne est de 18deg.C pendant l'hibernation profonde, soit approximativement 3 fois plus que la valeur moyenne et six fois plus que la valeur minimale des pertes des marmottes reproductrices.
Les premiers signes de début de la période de pré-reproduction de l'hibernation ont été enregistrés fin janvier chez certaines marmottes. Les testicules des mâles et les parties externes du vagin des femelles s'accroissent et un mucus blanc est souvent présent dans ou près du vagin. Simultanément, la température de ces marmottes s'élève en fluctuant pendant 2-3 semaines de 4 et 37deg.C. (Fig. 1a). La mise en route de la période de pré-reproduction diffère d'un couple d'animaux à l'autre, fin janvier chez les marmottes les plus précoces et fin février-début mars chez les plus tardives. Cependant, les membres d'un couple sont synchronisés comme ils le sont pour les réveils en cours d'hibernation.
Durant la période pré-reproductive, la tempé-rature corporelle est élevée et instable, les testicules et les parties vaginales externes continuent de se développer. Les frottis vaginaux de toutes les femelles reproductives montrent différentes phases d'oestrus (oestrus, dioestrus, métaoestrus et proestrus). A la fin de cette pé-riode (15-24 jours après), la température corporelle des marmottes reproductives atteint un niveau élevé et stable (36-38deg. C) et toutes les femelles reproductrices sont en oestrus (Fig. 1a) et les testicules atteignent un développement maximal, coïncidant avec le début de l'activité réelle des marmottes et de la période reproductrice dans un couple. Les pertes de poids sont maximum au cours de la période pré-reproductrice de l'hibernation. Cependant, leur dynamique varie suivant les couples. Les indices de perte de masse dépendent des moments du début de préparation à la reproduction. Habituellement, les moments de pertes pondérales moyennes masquent la situation réelle (Tab. 1). Par exemple, les pertes pondérales des marmottes, dont la période de pré-reproduction commence en janvier (1991-92), atteignent 5,0-5,6 g/kg par jour de la mi-janvier à la mi-février. Au même moment, les pertes pondérales des couples, commençant leur période pré-reproductive plus tard, sont seulement de 0,9-1,5 g/kg par jour comme pendant l'hibernation profonde. L'accouplement des couples reproducteurs commence après la fin de l'hibernation (niveau élevé et constant de la température corporelle). Pendant cette période, les dépenses énergétiques sont très importantes. Habituelle-ment, les animaux ne s'alimentent pas pendant toute cette période et présentent les pertes de poids les plus élevées. Certains animaux reproducteurs jeûnent tout au long de la gestation et même pendant la lactation. Chez les animaux reproducteurs, aucune chute de la température, d'arrêt de l'oestrus, de la gestation, de la lactation chez les femelles ni d'arrêt de l'activité sexuelle chez les mâles n'est enregistré même pour un poids inférieur à 3 kg. Avec une telle masse, les marmottes non reproduc-trices continuent à hiberner jusqu'au début de la période alimentaire. La longue période de jeûne (environ 40 j) et les basses températures (jusqu'à -20deg.C) observées pen-dant les périodes d'accouplement et de gestation ne provoquent pas de chute de la température corporelle. Les retours à l'hibernation n'ont été observés (fin d'avril) que chez deux femelles allaitantes dont la masse (2,2 et 2,3 kg) avait chutée de 50% entre février et avril. Chez ces dernières, l'arrêt de la lactation a précédé la baisse de la température corporelle. La dynamique de celle-ci et de la masse varient suivant les individus. Certains couples de marmottes non reproducteurs, mais qui ont eut une portée l'année précédente ou suivante, présen-tent la même évolution de température et de masse corporelles, les mêmes cycles oestriens chez les femelles et le même développement des testicules chez les mâles que celles de couples reproducteurs. L'absence de juvéniles dans ces familles dépend, probablement, de facteurs nonénergétiques.
Chez les marmottes, d'un poids de 3,4-3,7 kg au début de la période pré-reproductive, l'évolution de la température corporelle et du vagin est la même que celle des animaux reproducteurs (Fig. 1b). Leur température remonte fin janvier/mi-février aussi, mais celle-ci ne devient constante qu'après la prise alimentaire, c'est-à-dire que l'hibernation continue pendant cette période. Simultanément, aucun oestrus stable n'est observé chez les femelles (Fig. 1b) et les testicules des mâles ne sont, probablement, pas suffisamment développés. Les pertes de poids de ces marmottes ne sont que peu différentes (19,9%) de celles des marmottes reproductrices (20,6%). L'absence de nourriture entraîne une chute de leur température (Fig. 1b), ainsi que l'arrêt de l'oestrus et la réduction des testicules. Lorsque l'alimentation a débuté (mi-mars), leur température augmente et devient constante (début effectif de l'activité), les femelles sont en oestrus et les testicules des mâles développés. Mais ces couples n'ont jamais de descendance.
Les marmottes dont le poids est inférieur à 3,4 kg au cours de la pré-reproduction (mi-janvier) ont une température plus basse. Le pré-oestrus prédomine chez les femelles (Fig. 1c) et les testicules ne se développent pas chez les mâles. Ces animaux passent, à nouveau, en hibernation profonde après une courte période pré-reproductive à la fin de l'hiver. Cette dernière, chez les animaux encore plus légers, n'est jamais observée. Ils sont en hibernation profonde jusqu'au début de la prise alimentaire. Fin décembre 1987, deux de ces femelles ont présenté des indices de pré-reproduction. Cependant ni leurs cycles oestriens ni l'évolution de leur tempé-rature n'ont été enregistrés, mais par contre leurs pertes de poids entre septembre-décembre ont été à peu près les mêmes que celles des marmottes reproductrices entre septembre-janvier.
*****
Figure 1. Evolution de la température corporelle et du cycle oestrien. Dynamic of body temperature and estral cycle
Fig. 1A
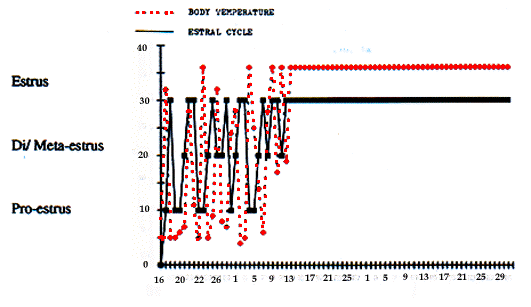
Fig 1B
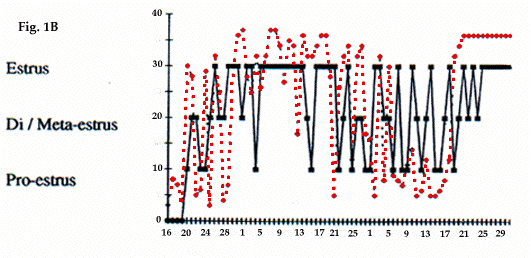
Fig 1C
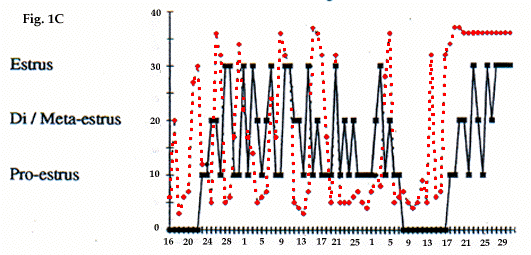
Tableau 1. Evolution de la masse corporelle des marmottes reproductrices entre
septembre et février 1990-91.
Table 1. The dynamic of body mass of breeding marmots in September-February
1990-91.
| Date | Marmot number | Body mass (kg)/Mass losses (g. per kg and per day) | |||||
| males | females | ||||||
| mean | maximum | minimum | mean | maximum | minimum | ||
| 20 Sept. | 56 | 7.2 / 0 | 9.1 / 0 | 5.5 / 0 | 6.1 / 0 | 8.7 / 0 | 4.9 / 0 |
| 01 Nov. | 56 | 6.3 / 3.3 | 7.7 / 4.2 | 4.7 / 2.2 | 5.2 / 3.7 | 7.5 / 4.6 | 4.4 / 2.3 |
| 10 Dec. | 30 | 5.7 / 1.4 | 7.1 / 1.9 | 4.2 / 0.6 | 4.8 / 1.0 | 7.0 / 1.7 | 3.8 / 0.5 |
| 15 Feb. | 22 | 5.2 / 2.5 | 6.8 / 5.6 | 6.6 / 1.1 | 4.4 / 2.4 | 6.6 / 5.3 | 3.3 / 0.9 |
The first period of real marmots hibernation in husbandry began in September after the end of feeding. The mean body mass of breeding steppe marmots at this time was 6.1 kg. in females and 7.2 in males. We did not registered breeding in animals which body mass in the middle of September was less than 4.9 kg. in this time (Tab. 1). Body mass losses of breeding marmots during the first period of hibernation (middle of September -beginning of November) were considerable (Mean losses: 13.5 % from body mass during the first 40 days of hibernation or 3.4 g/kg per day). The mean body temperature of breeding animals decreased in this period from 32 to 14deg. C. The minimal body mass of these marmots the 1st of November was 4.4 kg. (Tab. 1). Body mass losses of breeding marmots in the period of "deep" hibernation (November-middle of January) were minimal (1.3 g/kg/day). The minimal indexes of body masses were 3.8 kg for females and 4.1 kg for males in the middle of January (Tab. 1). Dynamics of body tem-perature were individual during the period of deep hibernation as well as arousal number and dates and mean body temperature. Both did not depend from am-bient temperature. Mean body temperature of the ani-mals was 9.7deg. C. Mass losses were more important in animals with a high mean body temperature and more frequent arousal. Thus mass losses of a unbreeding marmots, which mean temperature was 18deg. C through the "deep" hibernation, were 3.1 g/kg in day, i.e., approximately 3 times more than the mean of losses and 6 times more than the minimal ones of breeding marmots.
First signs of the onset of prebreeding period of hibernation were registered at the end of January in some marmots. Testicles in males and external parts of vagina in females increased. White mucous secretion often was found in and around vagina. Simultaneously body temperature of that marmots increased and fluctuated from 4 to 37deg. C. during 2-3 weeks (Fig 1a). Dates of prebreeding period onset differed between marmot pairs: end of January for early pairs and end of February-beginning of March for the latest ones. How-ever, within pairs this process was rather synchronical as the arousal during deep hibernation. During the period of high and unstable body temperature (pre-breeding period of hibernation) testicles and external part of vaginas continued to develop. Vagina smears showed different phases of oestral cycles (oestrus, di-oestrus, metaoestrus and pro-oestrus). At the end of this period (through 15-24 days after its beginning) body temperature of breeding marmots reached a high and constant level (36-38deg. C). Simultaneously in vagina smears of all breeding females there were only charac-teristic oestrus cells (Fig. 1a) and testicles of males reached their maximal growth. Beginning of these processes in a pair indicate the beginning of real activity and onset of breeding period. The losses of marmot body mass were maximum during prebreeding period of hibernation. However their dynamics were very different in animal pairs. The indexes of mass losses depended on the dates onset of preparing to breeding. As usual the dates of mean losses masked the real situation (Tab. 1). For example, the mass losses of marmots which prebreeding period began in January (1991-92) reached 5,.0-5.6 g/kg in a day from the middle of January to middle of February. At that time the losses of mass in pair, beginning prebreeding period later (end of February), were only 0.9-1.5 g/kg in a day like in the "deep" hibernation. coupling in pairs of breeding marmots began after termination of hiberna-tion (high and constant level of body temperature). During that period the energetic expenditure of mar-mots are very large. As usual, animals did not forage all that period and showed the greatest mass losses. Some breeding marmots were starving through all pregnancy and even part of lactation period. Any decrease of body temperature, stop of oestrus, end of pregnancy or lactation in females and stop of sexual activity in males had been registered even when the body masses of breeding marmots fall under 3 kg. But unbreeding marmots with the same body mass continued hibernation up to the beginning of feeding. The long starvation (about 40 days) and low ambient temperature (to -20deg. C) during coupling and pregnancy periods did not provoke a decrease of body temperature of breeding marmots. Recurrence of hibernation were observed only in two lactating females (end of April) which lost a conside-rable part of their body mass (about 50% from Februa-ry to April). For these very weak and light body mass (2.2 and 2.3 kg) females lactation stop preceded body temperature decrease. Body temperature and weight dynamics between autumn-spring were very variable. Some of non-breeding marmot pairs had the same body temperature and body mass dynamics, oestral cycles in females and development of testicles in males than the breeding ones. In some of these pairs, litters were regis-tered next or previous year. Probably lack of young in these families depended on non-energetic reasons.
Body temperature dynamics, vagina dynamics and testicles development of marmots with a light body weight at the onset of prebreeding period (3.4 - 3.7 kg) were similar to those of breeding animals (Fig. 1b). Increase of their temperature began at the end of January-middle of February too. However this tempera-ture never became constant before the onset of foraging, i.e., their hibernation continued for that time. Simulta-neously in females constant oestrus was not registered (Fig. 1b). Probably the testicles in male were not deve-loped enough. Losses of body mass of these marmots through hibernation (from September to the middle of January) were approximately the same (19.9 %) as in breeding animals (20.6 %). Lack of food provoked the decrease of body temperature of these marmots as well as oestrus and reduction of testicles. After onset of fora-ging (middle of March) their body temperature became high and constant (real onset of activity), females were constantly in oestrus and males had a high development of testicles. But these pairs never had litters.
Lighter marmots (less than 3,4 kg in the middle of January) in pre-breeding period had a smaller body temperature. In females pre-oestral phases predomi-nated and testicles were not developed in males. These animals often fall again in deep hibernation after short prebreeding period at the end of winter For weaker marmots prebreeding period in the second part of winter was not registered at all. These marmots were in deep hibernation up to the beginning of foraging in spring. In 1987 two females from this group showed white vaginal secretion, indices of prebreeding, at the end of December. Unfortunately their oestral cycles and body temperature dynamics were not investigated but their weight looses in September-December were approximately the same of those of breeding marmots in the September-January.
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |