![]()
Deux groupes familiaux de marmotte alpine ont été suivi pendant
sept ans (1988-1994) par observation directe dans les Apennins d'Emilie. Ces
observations ont porté sur la démographie, l'organisation
sociale, l'extension du territoire familial, l'activité en
référence aux terriers et à l'utilisation du territoire.
Mots-clés : Marmotte alpine, Marmota marmota, Apennins, Italie, dynamique des
populations, territoire, terriers.
Two contiguous families of Alpine marmot were followed for 7 years
(1988-1994) by direct observation in the Emilian Apennines. Observations
regarded demographic development, social organization, extension of the family
territory, activity with reference to burrows and use of territory.
Key words: Alpine marmot, Marmota marmota, Apennines, Italy, population dynamics,
use of territory, burrow systems.

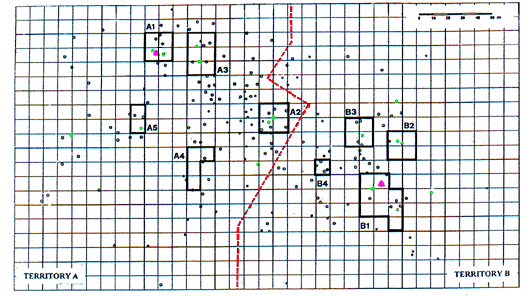
Les entrées des terriers du site ont été marquées par des jalons numérotés visibles de loin et leur examen topographique a été entrepris (Sala et al., 1992) en divisant l'aire étudiée suivant une grille de 10x10 m. Ces données ont été traitées à l'aide du logiciel AutoCad 1.0 (Microsoft), et l'aire fréquentée par les marmottes déterminée par la méthode du polygone convexe (Schoener 1981; in Mykytka & Pelton 1988).
A partir de 1988, les marmottes de ce site ont été marquées (Sala et al. 1993) et leur classe d'âge (marmottons, jeunes (1-2 ans), ou adulte) enregistrée suivant la classification d'Arnold (1990). Plus de 660 heures d'observations ont été réalisées pendant 66 jours de mai à septembre.
La présence, la position dans le site et l'activité de chaque animal ont été enregistrées toutes les 15 mi-nutes suivant la méthode d'échantillonnage par balayage (Altman 1974), ainsi que la distance au plus proche terrier. L'activité des marmottes a été catégorisée : 1. interne au terrier, 2. en mouvement, 3. assise, 4. bain de soleil, 5. état d'alerte, 6. dressée, 7. alimentation, 8. cri, 9. combat, 10. jeu, 11. fuite, 12. interaction amicale, 13. toilettage, 14. auto-toilettage, 15. chasser un congénère (non par jeu), 16. transport d'herbe, 17. momentanément non visible, 18. fouissement, 19. marquage odorant, 20. repos allongée, 21. s'apprête à rentrer au terrier. Les activités individuelles ont été groupées en 6 classes : I. alimentation (7) ; II. repos (3, 4, 20) ; III. mouvement (2, 11, 16, 18, 21) ; IV. interaction (10, 12, 13) ; V. comportement d'alerte (5, 6, 8) ; VI. autres (14, 17, 19).
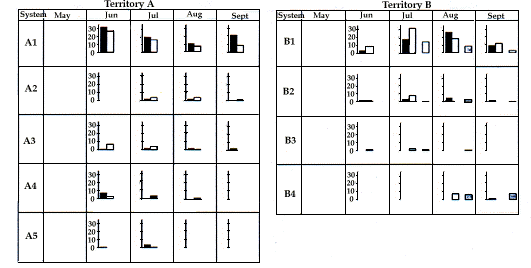
En B, le système principal est en position cen-trale, comme dans les Alpes (Perrin et al. 1992), l'hiber-nation et l'élevage des marmottons sont réalisés dans le système B1. Au contraire, en A, le système A1 est loca-lisé près de la bordure du territoire, alors que la femelle a utilisé les systèmes secondaires A2 (1991) et A4 (1992) pour l'élevage des marmottons et que l'hiberna-tion se passe dans le système A4. Ainsi, comme cela a été confirmé par l'observation de deux autres familles voisines, l'utilisation de différents systèmes pour l'hiber-nation et au cours de l'été ne semble pas être la règle dans les Apennins. Quand des terriers différents sont utilisés en été et en hiver, les derniers sont à une altitude plus basse dans les zones moins pentues où la neige s'accumule assurant une isolation thermique plus effica-ce pendant l'hiver. Une situation similaire a été rap-portée chez Marmota flaviventris (Svendsen 1976).
Dans les deux territoires, les systèmes principaux sont utilisés surtout pour le repos (plus de 80% en A; Fig. 4). D'un autre côté, l'alimentation y est moins fré-quente qu'autour des systèmes secondaires (Fig. 5). Le très faible pourcentage de comportement de surveillance dans le système principal (0,5% en moyenne) suggère que les animaux s'y sentent particulièrement en sécurité. La principale activité au niveau des systèmes secon-daires est aussi le repos, surtout en A3 (73%) et en B2 (68%), terriers situés sur les pentes escarpées à la limite supérieure des deux territoires. Pour les terriers situés plus bas, le temps passé en repos chute graduellement jusqu'à un minimum de 20% dans le système A5.
Au contraire, le temps consacré à l'alimentation augmente dans les zones plates, avec un minimum au niveau des terriers les plus hauts (3% en A3 et 8% en B2) et un maximum au niveau des terriers les plus bas (46% en A5 et 18% en B4). Probablement, les animaux préfèrent s'alimenter dans les zones les moins pentues, alors que celles situées sur les pentes rocheuses, plus haut, offrant une meilleure visibilité et un meilleur contrôle du territoire sont choisies pour le repos.
Les forts pourcentages de déplacements autour d'A4 et de B4 peuvent s'expliquer par la proximité de ces systèmes à un sentier reliant les terriers des zones plates à ceux des zones supérieures.
Un pourcentage élevé d'interactions sociales est enregistré autour d'A2, A5 et B3 qui sont utilisés principalement par les jeunes et les un-ans.
Les comportements de surveillance, toujours plus nombreux à côté des systèmes secondaires que des systèmes principaux, atteignent 13,5% en A alors qu'ils n'excèdent pas 3% en B.
A proximité des terriers dispersés, le repos reste l'activité dominante (moyenne 29,5%), quoique moins importante que dans les systèmes principaux (67%) et secondaires (52%) (Fig. 6). Au contraire, l'alimentation (25,5%) et les déplacements (26%) augmentent. La fréquence des comportements de surveillance (2,3% en A et 7% en B) se situe entre celle du système principal (0,5%) et celles des systèmes secondaires (8%).
La surface des territoires familiaux est compara-ble à celle des Alpes et ne change pas de façon substantielle entre 1989 et 1992 en dépit de variations importantes de la taille et de la structure des familles.
La détermination quantitative de l'utilisation de chaque terrier montre l'existence de trois catégories : un système principal, des systèmes secondaires et des terriers dispersés sur le territoire.
Le système principal se distingue des autres par le fait que les animaux y consacrent plus de temps en repos, mais cette activité est cependant prédominante dans les autres systèmes où elle s'accompagne d'une fré-quence élevée d'alimentation. Celle-ci augmente gradu-ellement, au fur et à mesure que le repos diminue, des terriers les plus élevés sur les pentes les plus fortes aux terriers les plus bas et sur les zones les plus plates.
Quand la reproduction et l'hibernation se passent dans le même terrier (B1), c'est dans le terrier principal. Alors que lorsque différents sites sont utilisés pour la reproduction et l'hibernation, ce sont des terriers des systèmes secondaires (A2 et A4).
L'acquisition continue d'information sur la composition et la structure des unités familiales serait nécessaire pour clarifier les aspects démographiques.
*****
| Fig. 1. Individus résidant dans l'aire d'étude de 1988 à
1994. Les différentes tailles correspondent aux différentes
classes d'âges : adultes, jeunes (1 et 2-ans), et marmottons. Noir :
mâle, Blanc : femelle, gris : non identifié. Resident individuals in the study area from 1988 to 1994. The different sizes indicate the different age classes: adults, young (1 and 2 years old) and infants. Black, male; white, female; grey, unidentified. 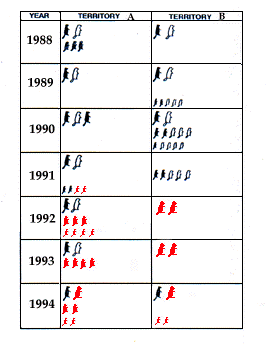
| Fig. 2. Fréquence moyenne des passages des frontières du
territoire familial par classe d'âge (1989-1992). Mean frequency (%) of crossing family territory boundaries by age class (1989-1992). 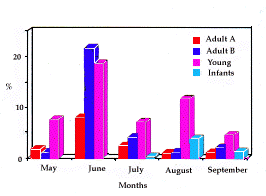
|
| Fig. 4. Fréquence moyenne (%) des différentes
activités dans les systèmes principaux de terriers Mean frequency (%) of the different activities in the main burrow systems. 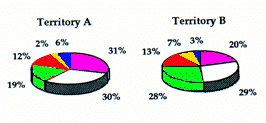
| Fig. 5. Fréquence moyenne (%) des différentes activités
dans les systèmes de terriers secondaires. Mean frequency (%) of the different activities in the secondary burrow systems. 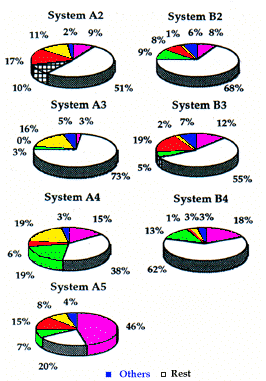
|
| Fig. 6. Fréquence moyenne (%) des différentes activités
dans les terriers dispersés. Mean frequency (%) of the different activities in scattered burrows. 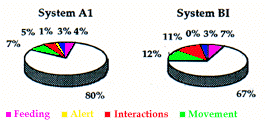
|
La fréquence moyenne des observations à chaque système de terriers est représentée par les polygones. Noir : mâle, blanc : femelle, gris : marmottons. / The average frequency of observations at the different burrow systems is shown by the polygons. Black: male, white: female, grey: infant.
| Fig. 9. Utilisation du territoire (family B) lors d'une année de
reproduction où les trois classes d'âge sont présentes. Use of the territory (family Bdeg. in a reproductive year in which the three age classes are contemporaneaoulsy present. | Fig. 10. Utilisation du territoire (famille A) lors d'une année de
reproduction.. Use of territory (family A) in a reproductive year. |
Scientific and management interests in this rodent in environments other than its original alpine habitat prompted a study to compare the structure and dynamics of the Apennine population with those in the Alps and to collect data on the use of territory.
Burrow entrances in the area were marked with numbered stakes which could be read from a distance, and a topographic survey of the location of the individual burrows was undertaken (Sala et al. 1992). The study area was divided up into a grid of 10x10 m. The topographic data were processed by computer using AutoCad 1.0 (Microsoft), and the area frequented by the marmots determined by the convex polygon method (Schoener 1981; in Mykytka & Pelton 1988).
From 1988 on, the marmots residing in this area were marked (Sala et al. 1993), and their age class [infant, young (1-2 years) or adult] recorded following Arnold (1990) classification. In all, more than 660 hours of observation were effected in 66 days, from May to September.
The presence, position in the area and the activity of each animal were recorded every 15 minutes using the 'Scanning Sample Method' (Altman 1974). Position was recorded as the distance from the nearest burrow, while activity was defined in terms of the following categories: 1. marmot in the burrow; 2. in movement; 3. sedentary; 4. sunbathing; 5. state of alert; 6. standing upright; 7. feeding; 8. calling out; 9. fighting; 10. playing; 11. running away; 12. friendly interaction; 13. grooming; 14. auto-grooming; 15. chasing a co-specific (not in play); 16. carrying grass; 17. temporarily hidden; 18. burrowing; 19 marking with scent; 20. recumbent rest; 21. about to enter burrow. Data were processed by grouping the individual activities into six classes: I. feeding (7); II. rest (3, 4, 20); III. movement (2, 11, 16, 18, 21); IV. interaction (10, 12, 13); V. alert behaviour (5, 6, 8); VI. other activities (14, 17, 19).
The number of members in each family varied strongly, both as a result of inconstant reproduction over the years (Fig. 1) and of high mortality rate in the family in territory B in 1991. It is known that in Alpine marmot reproduction and non-reproduction periods, alternate. However, the data reported by various authors differ strongly. According to Arnold (1990), reproduction occurs every other year in the Alps. Mann & Janeau (1988), Lenti Boero (1991) and Coulon et al. (1993) report two to five years of reproduction followed by a break of at least one year in the same group, while Nogué & Arthur (1992) have observed seven consecu-tive years of normal reproduction in four pairs in the Pyrenees. Allainé (1992) and Perrin et al. (1993) report reproductive periods not exceeding two years followed by a break of at least one year, the finding which corresponds most closely with our results: in each family, 3 reproductions in 7 years. For this reason, although the average size of the litter (3.66 animals) is in line with the highest values reported in the Alps (2.3 by Mann & Janeau, 1988; 4.1 by Lenti Boero, 1991; 3.1 by Nogué & Arthur, 1992), the average number of infants per family (1.57) is considerably lower than that reported by Mann & Janeau (2.6, 1988), Lenti (2.27; 1991) and Nogué & Arthur (2.4-4;1992), if not dissimilar to Allainé's figure (1.36; 1992).
Among the other entrances, selecting those where activity exceeds 2% and grouping them with nearby entrances, even if the latter are less used, secondary centres of activity (secondary burrow systems) can be observed (in decreasing order of use in each territory: A2, A3, A4, A5 and B2, B3, B4) (Fig. 3). In addition, there are numerous entrances scattered across the area which are only used occasionally.
In territory B, the main burrow is in a central position, as also observed by Perrin et al. (1992) in the Alps, and both hibernation and rearing of the offspring occur in the main system B1. On the other hand, in territory A, the A1 system is located close to the edge of the territory, while the female used the secondary system A2 (1991) and A4 (1992) for the rearing of off-springs. Hibernation also occurred in the system A4. Thus, as also confirmed by observation of two families in adjoining areas, the use of different systems for hibernation and during the summer does not seem to be the rule in the Apennines. When different burrows are used in the summer and winter, the latter are at lower altitude in the less steep areas where accumulation of snow is greater and thermal insulation during the winter more efficacious. A similar situation has been reported in Marmota flaviventris (Svendsen, 1976).
In both territories, the main burrow systems are mainly used for rest (up to 80% in A) (Fig. 4). On the other hand, feeding is less frequent in comparison to the secondary systems (Fig. 5). The extremely low percent-age of alert behaviour (on average 0,5%) suggests that the animals feel particularly safe in the main system.
The main activity in the secondary burrow systems is also rest, particularly in A3 (73%) and B2 (68%) which are on steep slopes at the upper extremes of the two territories. Descending towards the lower burrows, time spent in rest falls gradually reaching a minimum of 20% at system A5.
In contrast, time spent in feeding increases closer to the plane, rising from the minimum values recorded at the highest burrows (3% in A3 and 8% in B2) to the maximum levels at the lowest sites (46% in A5 and 18% in B4). Probably the animals prefer to feed at the less steep locations, while the higher sites on rocky slopes affording greater visibility and control of the territory are chosen for rest.
The high percentage of movement around A4 and B4 can be explained by the fact that these are located along the paths which connect the plane to the higher burrows.
A high percentage of social interaction is recorded around A2, A5 and B3 as these are mainly used by the young and yearlings.
The alert behaviour in the secondary systems is always higher than in the main systems, however there is considerably discrepancy in the two territories. In territory A, the figure is 13.5%, while in territory B it does not exceed 3%.
In the burrows scattered across the territory, rest remains an important predominant activity (on average 29,5%), although to a lesser extent than in the main (67%) and secondary (52%) systems (Fig. 6). On the other hand, feeding (25,5%) and movement (26%) increase. The frequency of alert behaviour in the burrows scattered across the territory (2.3% in A and 7% in B) lies between that for the main (0.5%) and the secondary (8%) systems.
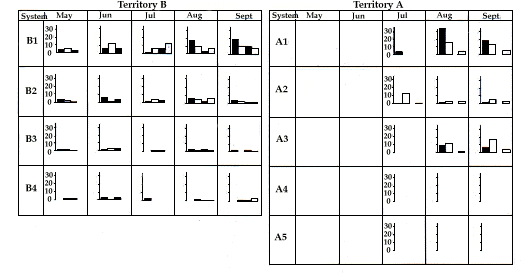
In the reproductive years, distribution of the activity of the adults does, however, differ in the two territories. In territory B, when only infants are present, the activity of the pair is centred on the area of the main burrow (Fig. 8). However, when the family embraces three generations (infants, young and adults), the activity of the female and the infants continues to be centred around the main burrow (which is also the site of reproductive burrow), while the male and the young move further afield, using, in particular, areas around the secondary burrow systems (Fig. 9). In contrast, in territory A, the activity of the male remains for the main part around the main burrow, while the female moves for reproduction to a secondary burrow, where she remains for some weeks after the emergence of the infants (Fig. 10).
In the first weeks after emerging from the burrow in which they were born, activity of the infants is centred only around this burrow, but from August and September their activity spreads across the territory to various burrows, particularly the secondary systems (Figs. 8, 9 et 10), in a pattern similar to that found in the one and two year old (Fig. 9).
The marmots observed in the area of study on Mount Cimone are divided into clearly defined families and territories. As in the Alps, reproduction in the Apennines is not annual, and the number of young recorded in a period of seven years (1988-1994) is particularly low.
The extension of family territories and the number of burrows is comparable with those found in the Alps and did not change substantially from 1989 to 1992 despite considerable changes in the size and structure of the resident families.
A quantitative assessment of the use of individual burrows revealed the presence of three categories: a main system, secondary systems, and burrows scattered across the territory.
The main burrow is distinguished from the secondary systems and scattered burrows by virtue of the fact that here the animals spend more time in rest, an activity which, however, remains predominant around the other categories, where it is accompanied by a higher frequency of feeding. This latter activity progressively rises, with the parallel fall in rest, when moving from the higher, steeper burrows to those in lower and flatter areas.
When reproduction and hibernation occur in the same burrow (B1), this is the main burrow, while if different sites are used, these are secondary systems (A2 and A4).
A continued acquisition of information on the composition and structure of the family units would therefore be welcomed in order to clarify demographic aspects further.
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |