Biodiversité chez les marmottes / Biodiversity in marmots,
Le Berre M., Ramousse R. & Le Guelte L. eds., International Marmot Network, 231-232. ISBN : 2-9509900-0-2
COCHET N., MEISTER R. & BARRÉ H.
Laboratoire Physiologie Genérale et Comparée, UCBL I - 43,
boulevard du 11 Novembre 1918 - 69622 VILLEURBANNE, FRANCE.
L'étude de la mobilisation saisonnière des
triglycérides contenus dans le tissu adipeux gonadique et
sous-cutané de marmottes alpines a révélé que sous
l'effet de facteurs stimulants, les adipocytes isolés de ces deux
dépôts montraient un taux lipolytique différent. Durant la
période de jeûne, le dépôt sous-cutané est
apparu plus lipolytique que le gonadique ce qui suggère qu'il pourrait
être le principal fournisseur d'énergie pendant l'hibernation.
Mots-clés : Marmotte alpine, tissu adipeux blanc, adipocytes,
lipolyse.
The study of the seasonal triacylglycerol (TG) mobilization from gonadal and
subcutaneous white adipose tissues of alpine marmots have revealed that during
the fasting period, isolated adipocytes from those two depots, stimulated with
different lipolytic factors, showed different lipolytic rates. The subcutaneous
depot appeared more lipolytic than the gonadal one and this suggests that this
depot would be the main energy supplier during the hibernation season.
Key-words: Alpine Marmot, white adipose tissue, adipocytes,
lipolysis.

En été, les marmottes accumulent des réserves de graisses
qui constituent leur seule source d'énergie pendant la période de
jeûne de l'hibernation. Les lipides sont stockées dans le tissu
adipeux blanc sous la forme de triglycérides (trois acides gras
estérifiés avec la fonction alcool du glycérol). Le taux
lipolytique, régulé par des hormones, consiste en une hydrolyse
des trigly-cérides en acides gras libres et en glycérol,
relâchés dans la circulation sanguine. Chez quelques
espèces de rongeurs, comme le rat et le hamster, une variation locale de
la lipolyse a été démontrée (Lacasa et al.
1991, Dieudonné et al. 1992), mais la mesure de la lipolyse sur
des adipocytes de différents dépôts n'a jamais
été réalisée au cours du cycle complet chez une
espèce hibernante. Le but de cette étude est d'évaluer la
contribution des dépôts adipeux gonadique et sous-cutané
dans la mobilisation lipidique au cours des périodes de jeûne
spontanée (hibernation) et d'alimen-tation des marmottes alpines.
Des fragments de tissus adipeux blanc de chacun des dépôts sont
prélevés chirurgicalement et les adipocytes isolés
préparés selon la méthode de Rodbell (1964). Les cellules
en suspension dans le liquide tampon de Krebs sont incubées avec une
forte concen-tration de norépinéphrine (10-5 M), d'ACTH (10-7 M)
ainsi qu'avec de l'Adénosine déaminase (ADA, 2 ug/ml) qui
inactive l'adénosine ambiante, inhibiteur local et physiologique bien
connue de la lipolyse basale. Les taux lipolytiques sont mesurés par
quantification du relargage du glycérol dans le milieu et
exprimés en umol / 100 mg de cellules / heure d'incubation à
37deg.C.
Pour les trois facteurs stimulants utilisés, le taux lipolytique des
adipocytes sous-cutanés est plus élevé que celui des
cellules gonadiques au cours de la période de jeûne. Ce
phénomène semble s'inverser au cours des périodes
d'alimentation, suggérant que les marmottes peuvent mobiliser
préférentiellement l'éner-gie d'un dépôt
particulier suivant la saison. Les résultats concernant la
période d'alimentation apparaissent en accord avec ceux mis en
évidence chez le rat et le hamster (Lacasa et al. 1991,
Dieudonné et al. 1992). Cependant, les résultats obtenus
au cours de l'hibernation montrent que, contrairement au hamster (Bartness
et al. 1989), les marmottes, hibernants de grande taille, mobilisent
essentiellement les réserves de graisse sous-cutanée plutôt
que celles d'origine gonadique. Cette observation confirme que les
dépôts sous-cutanés de graisse ne jouent pas un rôle
de protection thermique pour les marmottes (Webb & Schnabel 1983), mais
constituent une réserve importante d'énergie pour survivre
à l'hiver.
Ces résultats préliminaires suggèrent que le tissu
adi-peux sous-cutané pourrait être le principal fournisseur
d'énergie au cours de l'hibernation jusqu'au début de la
période estivale et que l'adénosine serait un modulateur possible
de la lipolyse des adipocytes blancs chez la marmotte.
*****
Figure 1. Lipolyse saisonnière d'adipocytes isolés
gonadique (barres) ou sous-cutané (O) de marmotte. / Seasonal
lipolisis of marmot isolated adipocytes from gonadal ( bars ) and subcutaneous
(O) fat depots.
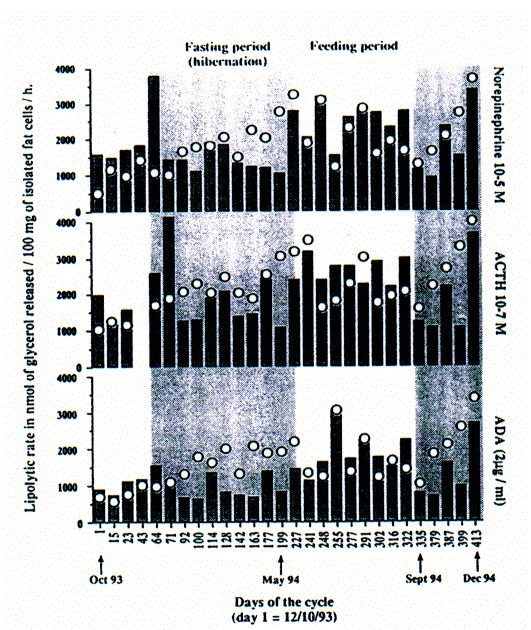
Stimulation par norépinéphrine 10-5M, ACTH 10-7 M et ADA
(2 ug/ml). Chaque couple baton-cercle correspond aux données d'un
animal différent. Début du cycle octobre 1993, fin en
décembre 1994. La période spontanée de jeûne des
marmottes est indiquée par une bande grise. / Stimulated by
norepinephrine 10-5 M, ACTH 10-7 M and ADA (2 ug/ml). Each couple stick-circle
corresponds to data from one different animal. The cycle began in October 1993
and ended in December 1994. Spontaneous fasting period of marmots are indicated
by grey bands.
In summer, Alpine marmots store fat its only energy source during the
hibernation fasting period. Fat is stored in white adipose tissue in the form
of TG (3 fatty acids esterified with the alcohol functions of glycerol).
Lipolytic rate, regulated by hormones, consist in an hydrolysis of TG into free
fatty acids and glycerol which are released in the circulating blood. In some
rodent species, like rat or hamster, regional variation of lipolysis have been
demonstrated (Lacasa et al. 1991, Dieudonné et al.
1992) but lipolytic measurements on isolated adipocytes from different
depots have never been realised during a complete cycle of an hibernating
species. The aim of the study was thus, to evaluate the contribution of gonadal
and subcutaneous adipose depots in lipid mobilization during spontaneous
fasting (hibernation) and feeding periods of alpine marmots.
Isolated white adipocytes from each depot, surgically removed from captive
animals, were prepared according to the Rodbell method (1964). Cells suspended
in a Krebs buffer were incubated with high concentration of norepinephrine
(10-5 M), ACTH (10-7 M) and also with Adenosine Deaminase (ADA, 2ug/ml) which
inactivates ambient adenosine, a well known local and physiological inhibitor
of basal lipolysis. Lipolytic rates were measured by quantification of the
glycerol released in the medium and expressed in umol /100 mg of cells / hour
of incubation at 37deg.C.
For the 3 factors used, the lipolytic rate of sub-cutaneous adipocytes was
higher than that of gonadal cells during the fasting period. This phenomenon
seemed to be reversed during feeding periods, sugges-ting that marmots can
mobilize preferentially energy from particular depots according to the season.
Results concerning feeding period appear in accordance with those found in rat
and hamster (Lacasa et al. 1991, Dieudonné et al. 1922).
However, results obtained during hibernation showed that conversely to hamster
(Bartness et al. 1989), marmots, larger size hibernators, mobilize
rather subcutaneous than gonadal fat stores. This observation confirms that
subcutaneous fat depot does not play the role of an insulative protection for
marmots (Webb & Schnabel 1983) but constitutes an important energy storage
for winter survival.
These preliminary results suggest that the subcutaneous adipose tissue may
be the major energy supplier during hibernation season until the beginning of
summer period and that adenosine appears as a putative local modulator of
lipolysis of marmot white adipocytes.
LACASA D., AGLI B., PECQUERY R. & GUIDICELLI Y 1991. Influence of
ovariectomy and regional fat distribution on the membranous transducing system
controling lipolysis in rat fat cells. Endocrinology, 128:747-753.
DIEUDONNE M. N., PEQUERY R. AND GUIDICELLI Y 1992. Characteristics of the
[[alpha]]2/ß-adrenoceptor coupled adenylate-cyclase system and their
relationship with adrenergic responsiveness in hamster fat cells from different
anatomical sites. Eur. J. Biochem., 205: 867-873.
RODBELL M. 1964. Metabolism of isolated fat cell. I: Effects of hormones on
glucose metabolism and lipolysis. J. Biol. Chem., 239: 375-380.
BARTNESS J. T., HAMILTON J. M., WADE G. N. & GOLDMAN B. D. 1989. Regional
differences in fat pad response to short days in siberian hamster. Am. J.
Physiol. 257 (Regulatory Integrative Comp. Physiol. 26):
R1533-R1540,.
WEBB D. R. & SCHNABEL R. R. 1983. Function of fat in hibernators: thermal
aspects. J. Therm. Biol., 8 (4): 369-374.A suivre...
 retour ou sommaire
retour ou sommaire