![]()
En Yakoutie, 2 sous-espèces de marmotte sont présentes : M.
camtschatica bungei dans les monts Verkhoyan, Chersky et Momski (N-E) et
M. c. doppelmayeri sur les hauts du Vitim-Oliokminsky, dans le
Charo-Tokkinsky, et aux sources de l'Aldan, de Chulman et de Iengri
(chaîne de Zverev,) en populations indépendantes. Au cours des
temps historiques, l'aire de l'espèce était plus importante. Ces
60 dernières années, les populations de Kondakovsky (aval de
l'Indigirka) et celles des lacs Sutorokha et Ozhogino (région
d'Abuisky), et d'une partie importante des monts de Momsky ont disparu. Le
futur de M. camtschatica dépend de l'homme, de ses efforts de
protection des populations existantes, des actions de réintroduction.
Mots-clés : M. camtschatica, destruction,
réintroduction, Verkhoyan, Chersky, Oliokminsky, Sutorokha, Ozhigino,
Yakoutie.
In Yakutia two subspecies of the black-capped marmot are present : M. c.
bungei in Verkhoyan, Chersky and Momski mountains (North-East) and M. c.
doppelmayeri in Vitim-Oliokminsky highlands, Charo-Tokkinsky country, Aldan,
Chulman and Iengri river-heads (Zverev mountain) as independent populations. In
historical time the species area was broader. During the last sixty years the
independent Kondakovsky population (lower reaches of the Indigirka-river) had
disappeared, as well as the colonies near the lakes Sutorokha and Ozhogino
(Abuisky region) and on a large part of Momsky mountain range. M.
camtschatica future depends on man, his efforts on protection of existing
population, on considered actions on its reacclimatization.
Key-words: M. camtschatica, destruction, reintroduction,
Verkhoyan, Chersky, Oliokminsky, Sutorokha, Ozhigino, Yakutia

Deux sous-espèces vivent en Yakoutie : M. camtschatica bungei (nord-est de la Yakoutie, monts Verkhoyansk, Chersky, Momsky, Kharaulakh) et M. c. doppelmayeri dans les Monts de Vitim-Olekma, entre deux fleuves du Chara-Tokko, aux sources de l'Aldan, Chulman et Iengra, au sommet du Zverev (Kapitonov 1960, 1978, Tavroskyi 1971, Revin & Lyamkin 1986, Revin 1987, 1990).
4.2. Terriers. Il y a trois types de terriers : d'hibernation, d'été et de secours (Kapitonov 1963a, b, Vasiliev 1989). Habituellement, une famille dispose d'un terrier d'hibernation, de 1-5 terriers d'été, et d'envi-ron 10 terriers de secours. Le nombre d'entrées dépend de la nature du site. La largeur des chambres d'hiberna-tion est de 0,25-0,60 m (moyenne 0,47), sa profondeur de 0,8-1,2 m, sa hauteur de 0,45-0,64 m, leur nombre dans le même terrier est 2-4, généralement 2. La masse de la litière est de 9 à 12,5 kg. Un terrier d'hibernation recouvre 8-176 m2. Les entrées sont situées au tiers de la profondeur, soit 0,5 m. Le réseau de galeries atteint une longueur totale de 15-65 m jusqu'à plus de 100 m. Les terriers sont souvent construits sur des corniches plates ou peu pentues, orientées au sud ou à l'ouest. Une importante couche de terre fine et la nourriture sont les éléments indispensables à sa construction. Les terriers d'été et de secours sont surtout construits dans des fentes de rochers, rarement dans un sol fin. Les marmottes ne peuvent pas construire de terriers profonds du fait du permafrost. Mais les animaux peuvent maintenir la chambre d'hibernation à une température de + 1,5/+ 2deg.C en orientant leur terrier à l'abri du vent et en l'échauffant activement : épaississement du sol sur 40 cm, fermeture des entrées par 2 à 10 couches de terre, d'herbe et d'air, colmatage des fentes par un mélange de terre et d'herbe.
4.3. Régime et comportement alimentaire. Dans la nature, la marmotte à tête noire consomme à la fois les parties aériennes et souterraines des plantes, différents invertébrés et aussi des petits mammifères et des oiseaux (Kapitonov 1963 a, b). Au cours de la période active, son alimentation change fortement. Au printemps, après l'éveil, jusqu'en juin, la marmotte con-somme essentiellement des racines et des rhizomes de légumineuses, des tiges et des bourgeons de saule. En été (mi-juin à mi-août), sa ration comprend des fleurs de légumineuses, plus faiblement les parties vertes et les graines d'autres plantes. En automne, les céréales et les parties souterraines de plantes pluriannuelles dominent. La composition spécifique de sa ration est déterminée par les caractéristiques des plantes de son habitat. Dans le Verkhoyan central, les principaux habitats de mar-mottes correspondent aux toundras à arbrisseaux, à graminées variées et à cobresia. Dans le Karaulakh, sur 125 espèces de plantes trouvées dans les habitats de marmottes, 80 sont consommées (Kapitonov 1963 a, b). Les fleurs et les feuilles sont les parties préférées, alors que les tiges font partie de la fraction non consommée.
La consommation alimentaire augmente du prin-temps à l'été. La consommation journalière, mesurée en conditions expérimentales, est de 85,5-116,2 g de ma-tière sèche chez les femelles d'un-an, de 128,1-144,7 g chez un mâle adulte. Des données similaires ont été obtenues chez M. bobac (Seredneva & Nezgorov 1977).
L'activité de surface par beau temps atteint 18 heures (Kapitonov 1978). Par temps couvert, l'animal sort peu et pour de courtes durées. Les marmottes s'éloignent peu de leur terrier pour s'alimenter, entre 2-3 à 50 m. Au cours des déplacements vers des zones plus éloignées, elles utilisent des terriers de secours. Entre les différents terriers, elles tracent des sentiers, qui peu-vent être profonds près du terrier principal. Les animaux adultes sont plus prudents que les jeunes. A l'approche d'un danger, ils émettent un cri d'alarme et les jeunes se précipitent dans les terriers, poussés par les adultes.
4.4. Reproduction. L'accouplement n'a jamais été directement observé. On suppose qu'il se produit au cours d'un des réveils printaniers, après quoi les ani-maux retombent dans un état de torpeur sans chute importante de leur température corporelle. La gestation dure 30-40 jours et les jeunes sont tous nés mi-juin (Kapitonov 1978), ce qui est confirmé par les marmottes de Baguzin (Zharov 1975). L'émergence des premiers marmottons s'observe lors de la première décade de juillet dans le Verkhoyan central. Apparemment les couples se forment en été, avant l'entrée en hibernation. La taille d'une portée de M. camtschatica bungei en Yakoutie est de 3-11 individus (moyenne 6, Kapitonov 1963 b, 1978), supérieure à celle de Barguzin (Zharov 1975) et celle du plateau de Koryak (Portenko et al. 1963).
4.5. Structure des populations. La sex-ratio des populations de marmottes présente une prédominance de femelles, 53-58% (Kapitonov 1978, Yakovlev & Shadrina, sous presse), caractéristique de cette espèce. Ainsi, dans les montagnes de Barguzin, 54-63% des femelles n'ont pas atteint l'âge de 3 ans et la sex-ratio ne s'égalise qu'à l'âge adulte.
La durée de vie maximum chez M. camtschatica bungei, sur le terrain, n'est pas inférieure à 6 ans (en captivité 9-11 ans). Malgré cela, la structure d'âge des populations du Karaulakh (Kapitonov 1978) et du Verkhoyan central (Yakovlev & Shadrina, sous presse) est caractérisée par une prédominance des marmottons. Une colonie de 8-23 individus est constituée de 1-5 familles, et 2-8 individus par famille (moyenne 4-5 ; Kapitonov 1978, Yakovlev & Shadrina, sous presse). Dans la région d'Yana-Indigirka, les marmottes d'une famille (2 à 22 individus) entrent en hibernation ensembles (Kapitonov 1978), mais rarement celles de deux familles. Dans ce cas, 30-40 individus peuvent se trouver dans le même terrier (Karpukhin 1964).
4.6. Causes de mortalité. Un taux élevé de mortalité des marmottons est observé (environ 50% en automne). Dans les principaux habitats du nord-est de la Yakoutie, 2,8-2,9 marmottons survivent à la fin de l'été, alors que la fertilité moyenne des embryons normaux est de 6,0. Les raisons de cette mortalité ne sont pas connues. L'influence de facteurs complexes est postulée, le principal est la faible viabilité des marmottons du fait de la consanguinité dans les colonies isolées et du braconnage. Les épizooties dans les populations yakoutes de marmottes n'ont pas été observées.
4.7. Hibernation. Ces marmottes ont la plus longue période d'hibernation de toutes les espèces de marmottes, même dans les zones à climat plus clément (Réserve naturelle de Barguzin). Elles hibernent 7,5-8 mois (d'octobre à mi-mai ; Svatosh 1926, Novikov 1937, Filonov 1961) et environ 9 mois dans les parties nord (septembre à mi-mai ; Kapitonov 1960, 1963). En captivité, l'hibernation dure longtemps, mais l'entrée en hibernation est retardée (fin octobre-début novembre) et la sortie avancée (durée d'hibernation expérimentale maximale : 1er novembre au 4 juin). Comme chez tous les hibernants, la période de transition avant l'entrée en hibernation profonde dure environ 2 semaines. Une diminution de la température corporelle pendant la torpeur précédant l'hibernation avec une diminution de la température du cerveau tous les deux jours, mise en évidence chez Citlleus beechevi (Strumwasser 1955), a été observée chez la marmotte à tête noire. Sa tempé-rature corporelle, au cours de l'hibernation, est toujours inférieure à 5deg. C. Un gradient de température décrois-sant des parties antérieures aux postérieures existe, comparable à celui mis en évidence par Dubois (1896).
4.8. Dénombrement. Quatre-vingt mille indi-vidus ont été estimés en Yakoutie dans les années 50 (densité moyenne : 25 animaux par km2, Kapitonov 1978). Depuis 1984, la densité diminue partout, et approche maintenant 9 animaux par km2. Une diminu-tion du nombre de colonies anciennes a eu lieu. Bien que l'aire de distribution de M. camtschatica reste la même, un démembrement intensif et un isolement croissant des aires est en cours, indiquant une dégrada-tion des populations de marmottes en Yakoutie.
4.9. Maladies. Aucun cas d'épizootie de peste lié à la marmotte n'a été observé en Yakoutie. L'infestation des animaux par des endo ou des ectoparasites est rare. Une seule espèce d'endoparasite est connue, Citellina triraciata, trois espèces de puces, Oropsylla silantiewi, Ceratophyllus lunatus, C. advenarius; une espèce de tique, Haemogamasus ambulans, et une espèce de poux, Enderleinellus longucaphalus (Kapitonov 1978).
4.10. Ennemis, compétiteurs symbioses. Les ennemis sont Nyctea scandiaca, Aquila chrysaetus, Canis lupus, Vulpes vulpes, Alopex lagopus, Gulo gulo, Ursus arctos ; les compétiteurs, Alticola macrotis, Ochotona hyperborea, Lepus timidus, Ovis nivicola, Rangifer tarandus. Les marmottes vivent sans compétition avec Falco peregrinus, Buteo buteo, Buteo lagopus et les invertébrés (Kapitonov, 1978).
1. Limites de la distribution actuelle; 2. 1-5 individus pour 100
km2; 3. 6-14 ind. /100 km2; 4. 15-25 ind./ 100
km2; 5. colonies distinctes; colonies disparues. / 1. borders of
the modern area; 2. 1-5 individuals per 100 km2; 3. 6-14 ind./100
km2; 4. 15-25 ind./ 100 km2; 5. separate colonies; 6.
disappeared colonies.
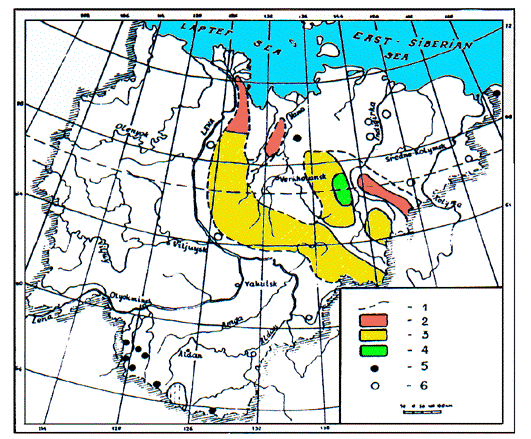
Two sub-species dwell in Yakutia: M. camt-schatica bungei in north-eastern Yakutia (Verkhoyansk, Chersky, Momsky, Kharaulakh mountain ranges), and M. c. doppelmayeri in Vitim-Olekma mount, in Chara-Tokko between two rivers, in river-heads of Aldan, Chulman and Iengra rivers, on Zverev mountain ridge (Kapitonov 1960, 1978, Tavroskyi 1971, Revin & Lyamkin 1986, Revin 1987, 1990).
4.2. Burrows. Three types of burrows are distin-guished: hibernation, summer and protection burrows (Kapitonov 1963 a, b, Vasiliev 1989). Usually a family has 1 hibernation, 1-5 summer and about 10 protection burrows. The number of entrance holes depends on site characters. Width of hibernation chamber is 0.25-0.60 m (average 0.47), its bottom is located 0.8-1.2 m under the surface, its height is 0.45-0.64 m, their number in one burrow is 2-4, usually 2. The weight of nest litter is from 9 to 12.5 kg. Hibernation burrows occupy 8-176 m2. The holes in them are located in one third of the depth near 0.5 m. Networks reach a total length of 15-65 m, in separate cases, more than 100 m. Hibernation burrows are constructed usually on terrace-like ledges, made on level plots or gentle slopes on southern or western sides. Presence of enough layer of fine earth and food are the important conditions for burrow construction. Summer and protection burrows are constructed mainly in cracks of rocks.
Marmots can't construct enough deep burrows in permafrost conditions. But, animals can maintain tem-perature in hibernation chamber on the level of + 1.5-2.0deg.C by locating burrow on lee side of slopes and also by active burrow warming: before hibernation they thic-ken the soil above hibernation chamber on about 40 cm, close the holes with 2-10 ground and grass and air layers, putty all cracks by mixture of ground and grass.
4.3. Diet and foraging behaviour.
In the field the black-capped marmots consumed both aboveground and underground parts of plants. Besides this, the animal eats different invertebrates and also small mammals and birds (Kapitonov 1963 a, b). During active period food composition changes considerably. In spring after arousal and up to June they consume mainly roots and rhizomes of Leguminous, stems and buds of willows. In summer (from middle of June to middle of August) the ration of marmots consists of flowers of Leguminous, in small degree of green parts and seeds of other plants. In autumn cereals and underground parts of perennial plants dominate in marmot ration. Plant species composition in the marmot ration is determined by character of vegetables of habitat. In Central Verkhoyan the main habitats of marmots are timed to shrub-motley grass-cereals-cobresia stepped tundra. In Kharaulakh mountains 125 plants species were found in marmots habitats, which eat about 80 species (Kapitonov 1963 a, b). Flowers and leaves are the most preferred parts. Stems of any plants are consumed.
Food consumption quantity increases from spring to autumn. Daily food consumption of 1 year-old females in experimental conditions was 85.5-116.2 g, in adult male: 128.1-144.7 g of dry food mass. Similar data were obtained for M. bobac which ate 108+/-6 g of dry mass (Seredneva & Nezgovorov 1977).
Marmots surface activity is higher in good wea-ther, when the duration outside activity is about 18 hours (Kapitonov 1978). In cloudy weather animals appear outside rarely and never for a long time. Mar-mots move not far away from burrows to forage: from 2-3 to 50 m. Otherwise they use protection burrows. Between burrows, as a rule, they tread paths, which turn into deep trench near the main burrow. Adult individuals are more careful than youngest ones. On seeing any danger, they utter the danger signal and young hide into burrows. In spite of this the adults literally push the gaping young into burrow.
4.4. Reproduction. Direct observations of the black-capped marmots coupling in nature are unknown. It is supposed that it occurs during one of the spring arousal, after which animals fall into a weak torpor without strong body temperature reduction. Pregnancy lasts 30-40 days and birth of juveniles is finished in the first half of June (Kapitonov 1978) which is confirmed by marmots from Barguzin mountains (Zharov 1975). The first appearance of juveniles in Central Verkhoyan is observed in the first decade of July. Apparently, future breeding pairs are formed before entrance into hibernation, in summer period. The size of litter in M. camtschatica bungei in Yakutia is equal to 3-11 individuals (average 6; Kapitonov 1963b, 1978), that is higher than on Barguzin mountains (Zharov 1975) and on Koryak plateau (Portenko et al. 1963).
4.5. Biological structure of population. Sex composition of marmot population is characterized by some predominance of females, 53-58% (Kapitonov 1978, Yakovlev & Shadrina, in print), which is obvious-ly peculiar for this species. So, on Barguzin mountains (Zharov 1975) 54-63% of females are under 3 years old and the sex ratio is equal only among adults.
Maximum duration of life in M. camtschatica bungei in natural conditions is not less than 6 year (in captivity: 9-11). In spite of this, age structure of popula-tions in Kharaulakh mountains (Kapitonov 1978) and Central Verkhoyan (Yakovlev & Shadrina, in print) is characterized by a predominance of juveniles. A colony consists of 1-5 families with commonly 8-23 animals. Moreover, the number of individuals in one family is 2-8 (average 4-5; Kapitonov 1978, Yakovlev & Shadrina, in print). According to observations conducted in Yana-Indigirka region, marmots of one family (2-22 animals) falls into hibernation usually together (Kapitonov 1978), rarely two families together. In the last case in one burrow 30-40 can be found (Karpukhin 1964).
5.6. Death causes. A very high death-rate among juveniles is observed (about 50% in autumn). So, in main habitats of north-eastern Yakutia 2.8-2.9 juveniles survive by the end of summer, whereas the average fertility of normal embryos is equal to 6.0. Reasons of death are not studied enough. The influence of complex factors is supposed, among which the main ones are low juveniles viability because of inbreeding in isolated colonies and also nonofficial hunting. Epizooties in marmots populations in Yakutia are not observed.
5.7. Hibernation. Black-capped marmots have the longer hibernation among all species of marmots even in regions with more comfortable climate (Barguzin nature reserve). They hibernate during 7.5-8 months (from October to middle of May; Svatosh 1926, Novikov 1937, Filonov 1961), in north parts hibernation goes on longer, about 9 months (from September to middle of May; Kapitonov 1960, 1963). Hibernation in captivity is also long, but entrance into hibernation starts later (end of October-beginning of November) and emergence sooner (maximum experimental hibernation duration was from November 1 to June 4).
As in all hibernating mammals, the transitional period before real hibernation lasts about 2 weeks. A body temperature reduction during sleepiness before hibernation was established by Strumwasser (1955), in Citellus beechevi. He observed the reduction of brain temperature every 2 days. We observed the same phenomenon in the black-capped marmots. Their body temperature during hibernation was always below 5deg.C, and front body part temperature is always higher. This gradient existence was found out by Dubois (1896).
4.8. Numbers. Marmots number in Yakutia in the 50ties was estimated as about 80 thousand individuals. Average density was 25 animals per 1000 km2 (Kapitonov 1978). Since 1984 population density falls everywhere and now is approximately equal to 9 mar-mots per 100 km2. Therefore, the present number of marmots is less than it was 50 years ago. A reduction of earlier known colonies number took place. Although, the area of M. camtschatica remains the same, an inten-sive crushing and an increasing isolation of areas go on, testifying of a gradual degradation of yakutian populations of marmots.
4.9. Diseases. Cases of mass murrains of mar-mots in Yakutia are not established. Animal infection by endo and ectoparasites is insignificant. Only one species of endoparasits is known, Citellina triraciata; three species of fleas, Oropsylla silantiewi, Cerato-phyllus lunatus, C. advenarius; one species of tick, Haemogamasus ambulans, one species of louse, Enderleinellus longucaphalus (Kapitonov 1978).
4.10. Enemies, competitors, symbiosis. The enemies are Nyctea scandiaca, Aquila chrysaetus, Canis lupus, Vulpes vulpes, Alopex lagopus, Gulo gulo, Ursus arctos; competitors: Alticola macrotis, Ochotona hyperborea, Lepus timidus, Ovis nivicola, Rangifer tarandus. Marmots can live together without competi-tion and enmity with Falco peregrinus, Buteo buteo, Buteo lagopus and Invetebrates (Kapitonov 1978).
BIBIKOV D.I. 1989. Marmots. Moscow, 258 pp.
BIBIKOV D.I. 1991. On taxonomy of genus Marmota In Biology, ecology
and rational use of marmots, Proc. of All-Union meeting, Seredneva T.A.
ed., Moscow: 16-19.
DUBOIS R. 1896. Physiologie comparée de la marmotte. Masson,
Paris.
FILONOV K.P. 196l. Materials on ecology of the black-capped marmot in Barguzin
nature reserve. In Workshops of Barguzin nature reserve, Moscow, 3:
169-180.
GROMOV I.M., BIBIKOV D.I., KALABUKHOV N.I., MEYER M.N. 1965. Surface sciuridae
(Marmotinae) The USSR fauna. Mammals. Moscow-Leningrad, 3(2): 466 p.
HOFFMANN R. 1968. Russian Science: a personal view. In: World Book
Encyclopedia, Science year: 228-239.
KAPITONOV V. I. 1960. The distribution of the black-capped marmot (Marmota
camtschatica Pall.). Bull. Mosc. soc. Nat. Testers, 65(5): 5-15.
KAPITONOV V. I. 1963. The black-capped marmot ecology and perspectives of
its practical use. Thesis of Dissertation for candidate's degree, Alma-Ata,
23 pp.
KAPITONOV V. I. 1966. Marmots distribution in Central Kazakhstan and
perspectives of their trade. In: Workshops of Zoological Institute of
Kazkhstan Academy of sciences, 26: 94- l 34 .
KAPITONOV V. I. 1978. The black-capped marmots. In: Marmots. Distribution
and ecology, Moscow: 178-209.
KARPUKHIN I. P. 1964. The black-capped marmot in north-eastern Yakutia. In:
Agricultural industry of siberia and far East, 5: 85-86.
KRIVOSHEEV V.G. 1988. Problems of teriography of North-eastern Asia. In
Common and regional teriography, Moscow: 33 - 74.
LUKOVTSEV YU. S., PERFILIEV V. I., YAKOVLEV F. G., VASILIEV V.N. 1990. The
black-capped marmot in Ust-Lena nature reserve. In: Resources of Siberia
fauna, Novosibirsk: 26l-263.
LYAPUNOVA E. A., BOYESKOROV G. G. &. VORONTSOV N. N. 1992. Marmota
camtschatica Pall. Nearctic element in palearctic Marmota fauna. In
Proc. 1st Intern. symp. on Alpine marmot, Bassano B., Durio P., Gallo
Orsi U., Macchi E., eds., Torino: 185-186.
NOVIKOV G.A. 1937. Hunting fauna of North-Western Baykal region. In:
Buryat-Mongolia, .M.-L., Izd.-vo AN SSSR.
REVIN Yu. V. 1987. Marmota camtschatica doppelmayeri
(south-yakutian population). In The Red Book of Yakutia, Novosibirsk,
13-14.
REVIN VU.V., LYAMKIN V.F. 1987. On area structure of the black-capped marmot
(Marmota camtschatica Pall. ) in near Baykal, north part of Baykal and
south Yakutia. In: Ecology and protection of mountains mammals species,
Proc. 3rd All-Union Conference, Moscow: 144-146.
SEREDNEVA T. A. & NEZGOVOROV A. L. 1977. Food consumption and processing by
steppe marmot (Marmota bobac). Zool. J., 56(12): 1839-1846.
STRUMWASSER F. 1959. Thermoregulatory, brain and behavioral mechanisms during
entrance into hibernation in the ground squirrels. Amer. J. Physiology,
196 (1): 8-14.
SVATOSH Z.F. 1926. Tarbagan. In Sable hunting in the north-eactern shore of
Baykal, Verkhneudinsk-Leningrad: 172-176.
TAVROVSKY V.A. ed. 1971. Mammals of Yakutia. Moscow, 660 pp
VASILIEV V.N. 1989. Hibernation ecology of the black-capped marmot (Marmota
camtschatica Pallas, 1811). Thesis of Disssertation for candidate's degree,
Moscow, 22 pp.
VERESCHAGIN N.K. 1959. Mammals remainders of mamoth epoch from taymir
Penincula. Bull. M. soc. Nature Testers, 64(5).
ZHAROV V. R. 1975. The black-capped marmot (Marmota camtschatica)
ecology on Barguzin mountain range. Thesis of Dissertation for candidate's
degree, Irkutsk, 24 pp.
ZHELEZNOV N.K. 1981. Rare mammals species of Chukotka autonomous area. In
Biological aspects of rare animals species protection, Moscow: 84-85.
ZHELEZNOV N.K. 1991. The characterization of the black-capped marmot
distribution. In Marmot populations structure, Moscow: 275-289.
ZIMINA R.P. & GERASIMOV I.P. 1971. Periglacial expantion of marmots in the
Middle Europe during upper pleistocene. Bul. Moscow soc. of Nature
Testers, 1: 7-15.
ZIMINA R.P. & GERASIMOV I.P. 1980. The History of genus Marmota and
the periglacial conditions importance of glacial period in its formation and
distribution. In Marmots. Biocenotic and practical importance, Moscow:
5-24.
A suivre
Forward![]()
| Retour à Documents scientifiques du réseau | Back to Scientific publications of the network |
| sommaire/contents |